DEZVOLTAREA NORMALA A EMBRIONULUI DE GAINA IN PRIMELE 72 ORE DE INCUBATIE
1. SEGMENTATIA
Segmentatia este procesul de transformare a zigotului unicelular in embrionul pluricelular si consta intr-o serie de diviziuni mitotice ce impart zigotul in blastomere.
Ovulul fertilizat este morfologic o singura celula cu un singur nucleu- | nucleu de segmentatie. Protoplasma vie este agregata in blastodisc, dar ramasitele ovulului sunt o masa inerta de materiale nutritive destinate spre a fi asimilate de catre embrionul care se ridica din blastodisc. In timpul diviziunilor timpurii, care se petrec sincronic in toate celulele, apare tendinta ca numarul celulelor sa creasca in progresie geometrica, ca mai apoi in timpul diviziunilor ulteriore aceasta sincronizare sa inceteze.
Controlul spatial (localizarea specifica a blastomerelor) si temporal (rata diviziunilor celulare) este realizat de ARN si proteinele citoplasmatice de origine materna ale zigotului (genomul zigotic nu este activ in fazele initiale ale segmentatiei astfel incat aceasta nu este afectata de inhibatorii transcriptiei ). Procesul este insotit de o scadere a raportului nucleo-citoplasmatic. Aceasta caracteristica se datoreaza abolirii perioadei de crestere intermitotica . pe de o parte si ratei foarte ridicate a diviziunilor nucleare , pe de alta parte. Rata diviziunilor nucleare o depaseste pe cea observata in celulele tumorale.
Transcriptia genomului embrionar incepe mai tarziu si reflecta probabil un anumit raport nucleo-citoplasmatic .Acum toate diviziunile sunt sincromizate. Tipul de segmentatie este influentat de :
cantitatea si distributia vitelusului in citoplasma;
factorii citoplasmatici cu rol in determinarea fimin - ului si pozitiei spatiale a fusului de diviziune.
In functie de distributia vitelusului in citoplasma zigotului . putem deosebi un pol celular bogat in vitelus ( polul vegetal ) si unul sarac in vitelus (polul animal) . unde se gaseste si nucleul , de cele mai multe ori [1,5] .
Zigotul cu o cantitate redusa (izolecit) sau medie (mezolecit) de vitelus prezinta o segmentatie de tip holoblastic sau totala , deoarece in urma omogenizarii vitelusului cu protoplasa . santurile de diviziune impart intregul zigot (Amphyoxus. Petromysontide , Ganoide , dipnoi , amfibieni . marsupiale plancentare).
Zigotul ce contine o cantitate mare de vitelus (telolecit sau centrolecit ) preyinta o segmentatie de tip meroplastic pentru ca vitelusul nu este bine penetrat de protoplasma limitand planul de diviziune cu formarea blastodiscului (Myxinoidee, teleosteeni , selacieni, reptile pasari, monotreme). La ouale telolecite segmentatia meroblastica este de tip discoidal, iar in cazul oualor centrolecite este de tip superficial.
La unele specii tipul de segmentatie depinde de factori externi : la Daphnia si altele nevertebrate inrudite oul este holoblast daca este produs vara si meroblast iarna .
Oul heteropolar (animale bilaterale) prezinta o simetrie bilaterala inainte de segmentatie, adica unul din planele meridionale determina asezarea partilor anterioara si posterioara embrionare .impartind oul in partea dreapta si stanga a embrionului.
Vitelusul (galbenusul) reprezinta o adaptare evolutiva ce permite dezvoltarea embrionara in absenta unei surse nutritive extreme, el trebuind sa asigure dezvoltarea embrionului in conditiile in care aceasta nu parcurge stadiile larvare si nu se implanteaza in uterul matern.
Zigotul de pasare prezinta o segmentatie de tip meroblastic discoidal (ca la moluste, cefalopode, pesti, reptile) adica implica doar citoplasma activa situata pe galbenus. In oul presegmental blastodiscul are un diametru de 3mm si aspect biconvex in sectiune .
Prima diviziune poate fi observata la aproximativ trei ore de la fecundatie. Moment ce corespunde patrunderii zigotului in istm, cu formarea a doua blastomere. La scurt timp (20 de minute) apare al doilea sant de diviziune perpendicular pe primul si se separa partial blastomerele. Celulele sunt complet separate : comunicarea se realizeaza la nivelul partii inferioare a blastodiscului si a protoplasmei inelului periferic periblastic. Al treilea sant se formeaza paralel cu primul in uter. Marimea celor opt blastomere devine variabila in stadiul de 32 celule cand se disting blastomerele centrale si cele marginale incoplct separate.
La patru ore dupa a treia diviziune se atinge stadiul de 256 celule.
Din stadiul de opt celule in partea profunda a blastodiscului apare un spatiu orizontal, separand protoplasma celulelor centrale de galbenus, numit spatiul suhgerminal sau cavitatea subembrionara , mai bine evidentiata in stadiul de 32 celule. Aceasta reprezinta prima delimitare ventrala a celulelor centrale.
Practic, spatiul subgerminal reprezinta o zona de lichefiere progresiva a vitelusului sub actiunea enzimelor din blastomerele centrale supradiacente iara a ajunge insa la nivelul blastomerelor marginale.
Din stadiul de 32 celulu apar primele diviziuni orizontale (paralele cu suprafata) ale celulelor centrale; este momentul in care zona celulara centrala devine bistratificata (si pluristratificata ulterior).
Consecutiv diviziunilor circumferentiale, nucleu celulelor marginale ajung in protoplasma inelului periblastic formand o structura de tip sincitial numita periblast sincitial. O parte din nucleu periblastului migreaza centriped in protoplasma plansului cavitatii subgerminale. Cu exceptia zonei centrale, corespunzatoare nucleului lui Pander. Se formeaza astfel sincitiul subgerminal care reprezinta primordiul peretelui germinai.
Periplastul celular realizeaza jonctiunea dintre grupul de celule marginale si periblastul sincitial (extern).
In acest stadiu se pot distinge doua zone ale blastomerului : aria pellucida - grupul de celule centrale supradiacente ale blastomerului; aria opaca- grupul de celule marginale si periblastul celular (aflate in contact direct cu vitelusul datorita lipsei spatiului subgerminal la acest nivel)
Celulele mai bogate in vitelus din aria pellucida se aglomereaza spre marginea inferioara a blastomerului formand portiunea centrala a hipoblastului . iar celulele sarace in vitelus se orienteaza superficial formand epiplastul (epiteliul unistratificat) . Initial, celulele prehipoblastice nu sunt organizate epitelial : alinierea lor intr-un strat unicelular incepe posterior si progreseaza anterior.
Anumite celule din epiplast migreaza in cavitatea subgerminala , formand hipoblastul primar. La acurt timp , o patura de celule din marginea posterioara a blastomerului migreaza anterior (cranial) , formand hipoblastul secundar , care se va uni cu cel primar.
In momentul depunerii oului, blastomerul contine aproximativ 60 000 celule organizate in doua straturi: epiplastul ( superior) ;
hipoplastul (inferior), separat dc o cavitate blastocelul.
HIPOBLASTUL
Celulele hipoblastului desi nu participa direct la formarea noului organism, au un rol important .Hpoblastul induce formarea si directionarea liniei primitivre Patten B.M. (1957) a demonstrat ca indepartarea marginii posterioare a blastodiscului opreste formarea
liniei primitive, iar aria pellucida se dezvolta ca o masa tisulara dezorganizata. In acest caz indepartarea hipoblastului opreste dezvoltarea care poate fi reluata numai dupa ce epiblastul genereaza un nou hipoblast.
Celulele hipoblastului din partea postero - marginala a blastodiscului induc formarea liniei primitive si astfel inhiba aparitia hipoblastului in alte regiuni marginale.
EPIBLASTUL
Lpiblastul nu este un tesut omogen, nediferentiat, celulele lui avand un anumit grad de diferentiere inainte de formarea liniei primitive. Numai unele celule epiblastice care prezinta la suprafata forma sulfatata a acidului glucuronic. migreaza si patrund in blastocel. restul celulelor ramanand la nivelul epiblastului.
Patrunderea in balstocel modifica forma celuleLor epiblastice care se alungesc, apoi se aplatizeaza si migreaza cu ajutorul filopodiilor. Dupa formarea ectodermului.
celulele acestuia secreta N - acetilglucozamina , acid hialuronic in blastocel care invelesc suprafata celulelor ingresate impiedicand adeziunea lor. Acidul hialuronic in mediul apos isi mareste volumul de peste 1000 ori.
Miscarea celulara insa este coordonata probabil de reteaua de fibronectina din membrana bazala a epiblastului . Reteaua apare cu putin timp inaintea formarii liniei primitive si lipseste in regiunea acestei linii; celulele liniei primitive migreaza lateral de-a lungul membranei bazale [3].
GASTRULATIA
Gastrulatia este un proces complex de formare a embrionului trilaminar din cel bilaminar caracterizata prin particularitati la nivel biochimic si celular. Modificari
biochimice se observa in cresterea Q.R. de la 0,7 -> 1 (valoarea caracteristica oxidarii glucidelor) prin care se asigura necesarul energetic de O2 al celulelor in timpul migrarii. Gastrulatia implica mai multe tipuri de miscari celulare:
imaginar ea si migrarea - deplasarea unor celule singulare sau a unor paturi de celule schimband raporturi de vecinatate;
epibolia - migrarea unor paturi celulare , de regula , ectodermale in jurul unor straturi vecine mai profunde;
involutia - migrarea spre interior a unui strat celular extern, care a captusi marginea interna a paturii celulare ramase la suprafasta;
ingresia - migrarea celulelor in mod individual de la suprafata spre interiorul embrionului;
delaminarea - separarea unei paturi in doua straturi mai mult sau mai putin paralele:
convergenta - deplasarea a doua regiuni spre acelasi punct: extensia - transformarea unei regiuni spre acelasi punct;
Miscarile celulare au fost urmarile prin metoda colorantilor vitali si metoda
celulelor.
Metoda celulelor marker consta in inlocuirea celulelor epiblastice ale embrionului gazda cu celule epiblastice marcate la nivel de ADN cu timidina tritiata ale embrionului donor. Astfel se poate urmari cu precizie migrarea celulelor si rolul lor. observandu-se ca toate structurile embrionare si cea mai mare parte a membranelor axtraembrionare deriva din epiblast. Hipoblastul arc doar o contributie mica la formarea lor. totusi celulele hipoblastice au un rol esential in dezvoltarea embrionara precoce.
FAZELE GASTRULAT1EI
Gastrulatia cuprinde doua evenimente majore:
A. Formarea liniei primitive;
B. Regresia liniei primitive.
.A.FORMAREA LINIEI PRIMITIVE
Structura majora care marcheaza inceputul propriu-zise la reptile, pasari si mamifere este linia primitiva. Ea apare dupa 6-7 ore de incubatie sub forma unei ingrosari la formarea careia participa celulele regiunilor laterale a jumatatii posterioare epiblastice ce migreaza spre centrul ariei pellucide formand linia primitiva initiala. La 12 - 13 ore de incubatie aceasta se alungeste anterior si se ingusteaza simultan formand linia primitiva intermediara. Alungirea spre anterior a liniei primitive este insotita de miscarea craniala a celulelor hipoblastului secundar
La 18 - 19 ore de incubatie embrionul atinge stadiul de linie primitiva definita care ocupa 23 din lungimea ariei pellucide prefigurand axul antero - posterior al embrionului. In urma alongarii liniei primitive aria pellucida isi schimba forma trecand din forma initiala circulara intr-una piriforma.
Miscarea de convergenta a celulelor epiblstice spre linia primitiva este insotita de formarea santului liniei primitive , achivalentul blastoporului embrionului amfibian, reprezentand zona de patrundere a celulelor in blastocel.
La extremitatea anterioara a liniei primitive apare o ingrosare numita nodul primitiv sau nodul Hensan care prezinta o depresiune echivalenta buzei dorsale a blastoporului.
Primele celule care patrund in blastocel la nivelul liniei primitive sunt cele din care se formeaza intestinul anterior primitiv. Odata ajunse in blastocel ele migreaza cranial si in cele din urma vor inlocui celulele hipoblastice din portiunea anterioara a embrionului.
Urmeaza apoi patrunderea in blastocel prin nodul Hensen a celulelor epiblastice. care migreaza anterior, dar ramana intre epiblast si hipoblast formand mezenchinul cefalic si notocordul. Toate aceste celule ce migreaza cranial determina ridicarea regiunii mediane anterioare a epiblastului adiacent, formand astfel procesul cefalic.
in ansamblu, in urma regresiei continue a celulelor la nivelul blastocclului se pot surprinde doua orientari de migrare a acestora:
una profunda, spre hipoblast care va fi inlocuit pe linie mediana si impins spre margini: aceste celule stau la originea organelor embrionare endodermale si majoritatea membranelor axtraembrionare;
una omnidirectionala a celulelor laxe intre epi- si hipoblast din care se va forma mezodermul embrionar si restul membranelor axtraembrionare.
Dupa 22 ore de incubatie , majoritatea celulelor precursoare endodermale se afla in blastocel . pe cand celulele precursoare mezodermale inca migreaza spre interior o perioada de timp.
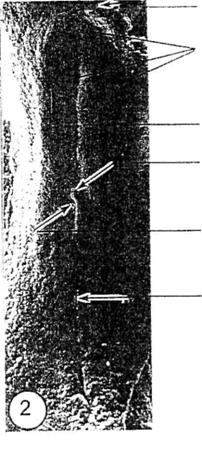
Descrierea embrionului la 18 ore de incubatie
Figura 1. Stadiu de linie primitiva definitiva (vedere dorsala)


|
Sant primitiv Figura Stadiu de pliu timpuriu cefalic (embrion intact) Piiu timpuriu ai capului |
|
|
Nod primitiv Cavitate primitiva
C. H. WADDINGTON a demonstrat rolul hipoblastului in orientarea axei embrionare. Mai tarziu AZAR si EYAE - GILADI au extins aceste observatii , aratand ca hipoblastul induce formarea liniei primitive. La indepartarea hipoblastului dezvoltarea este oprita aceasta reluandu-se doar dupa ce epiblastul genereaza un nou hipoblast. De asemenea, in cazul eliminarii marginii posterioare a blastodiscului, linia primitiva nu se formeaza , in aceasta situatie aria pellucida aparand ca o masa tisulara dezorgnizata.Deci . hipoblastul determina formarea si directionarea liniei primitive.
KHANER si EYAL - GILADI (1989) au demonstrat experimental ca locul de origine al celulelor hipoblastice este in partea postero - marginala a blastodiscului si ca aceasta inhiba formarea hipoblastului in alte regiuni marginale. Celulele zonei marginale prezinta un gradient de activitate, cu maximul la nivelul extremitatii posterioare.
ROLUL PRECURSORILOR EDODERMALI SI MI Ci RATIA ACESTORA
Celulele precursoare endodermale sunt localizate intr-o regiune ovala a epiblastului in jurul si inauntrul jumatatii craniale a liniei primitive initiale . Schoenwolf G.C.(1981) a sugerat ca epiblastul nu este un tesut omogen, nediferentiat asa cum s-a crezut pana acum. Se pare ca exista un grad de diferentiere a celulelor epiblastice inca inainte de formarea liniei primitive. Autorul a demonstrat ca numai unele celule epiblastice - cele care au o molecula de suprafata specifica ( o forma sulfatata a acidului glucuronic) - migreaza si patrund in blastocel , celelalte rampnand cantonate la nivelul epiblastului. 'Decizia' de migrare este luata deci Ia nivel celular si nu este o caracteristica a intregului eiplast.
Celulele migreaza spre hipoblast si incep sa-1 inlocuiasca craniolatcral. proces ce se deruleaza foarte rapid in stadiul de linie primitiva intermediara . Viitorul endoderm va fi localizat la capatul cranial al liniei primitive.
In stadiul de linie primitiva definita toti precursorii endodermali au ingresat iar hipoblastul inlocuit se condenseaza formand pe marginea cranoilaterala a ariei pellucide o zona in forma de semiluna.
in aceasta zona semilunara se diferentiaza celulele germina le primordiale care mai tarziu vor migra masiv si vor genera ovogoniile la femela si spermatogoniile la mascul [6].
ROLUL PRECURSURILOR MEZODERMALI SI MIGRATIA ACESTORA
Celulele viitorului mezoderm sunt localizate la capatul caudal al liniei primitive intermediare. O parte din ele vor forma mezodermul extraembrionar. Acesta este denumit astfel deoarece nu contribuie direct la formarea corpului embrionar, fiind precursorul notocordului si mezenchimul cefalic.
Ingresia precursorilor mezodermali are loc la capatul cranial al liniei primitive intermediare si migrarea se continua lent craniolateral (spre interior). Pentru mezenchinul cefalic si mezodermul inimii ingresia ia sfarsit in stadiul de proces cefalic , dar este continua pentru celulele partii craniale a notocordului.
Celulele viitorului mezoderm discoidal segmentat si discoidal lateral vor ingresa numai caudal . migrand de o parte si de alta a viitorului notocord. In schimb mezodermul plat si cel plat lateral se desfac intern [6].
ROLUL PRECURSORILOR ECTODERMALI SI MIGRATIA ACESTORA
In timpul gastrulatiei celulele precursoare ale ectodermului migreaza numai in bloc (prin epibolie) pentru a inconjura galbenusul.
Aceste celule nu migreaza sub forma de celule individuale ci in bloc. fiind unite prin jonctiuni stranse.
Numai celulele marginale ale ariei opace atasate strans de membrana vitelina. pe cand majoritatea celulelor blastodermului adera slab sau deloc. Spre deosebire de celelalte celule ale blastomerului, aceste marginale pot emite pseudopode foarte lungi (500 .) cu care se leaga de membrana vitelina. Prelungirile reprezinta aparatul locomotor al acestor celule. S-a dovedit experimental ca exista o recunoastere specifica intre celulele marginale ale ariei opaca si suprafata interna a membranei viteline care determina epibolia ectodermului. Probabil ca aceasta atasare este mediata de receptorii suprafetei celulare pentru fibroneclina de la nivelul suprafetei interne a anvelopei viteline
In stadiul de proces cefalic, ectodermul acopera mezenchimul cranial si mezodermul plat segmentat si se ingroasa formand placa neurala . In acest stadiu , mezodermul nu s-a
desfasurat in portiunea craniala a ariei pellucida. Aceasta zona se numeste proamnion si este formata numai din enctoderm si endoderm.
I.B.. REGRESIA LINIEI PRIMITIVE
Imediat dupa ce a atins lungimea maxima linia primitiva incepe sa regreseze astfel incat nodul Hensen se apropie de regiunea posterioara a ariei pellucide si atinge aceasta zona la 20 - 21 ore de incubatie. In acest moment epiblastul este constituit in intregime doar din celule ectodermale.
Datorita gastrulatiei bifazice apare un decalaj de dezvoltare la nivelul embrionului: in timp de portiunea posterioara a embrionului este inca in faza de gastrulatie. anterior a inceput organogeneza.
In timpul ingresiei celulelor precursoare ecto- si mezodermicc. celulele epiblastice precursoare celor ectoderme migreaza, pentru ca in final sa imbrace galbenusul prin epibolie in aproximativ patru zile. Astfel la sfarsitul gastrulatiei: ectodermul a inconjurat galbenusul in totalitate; endodermul a inlocuit hipoblastul; iar mezodermul este prezent intre ecto- si endoderm. Astfel s-a format embrionul trilaminat.
|
Politica de confidentialitate |
| Copyright ©
2024 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |