
ANATOMIA FUNCTIONALA A CEREBELULUI
A. SCOARTA CEREBELOASA
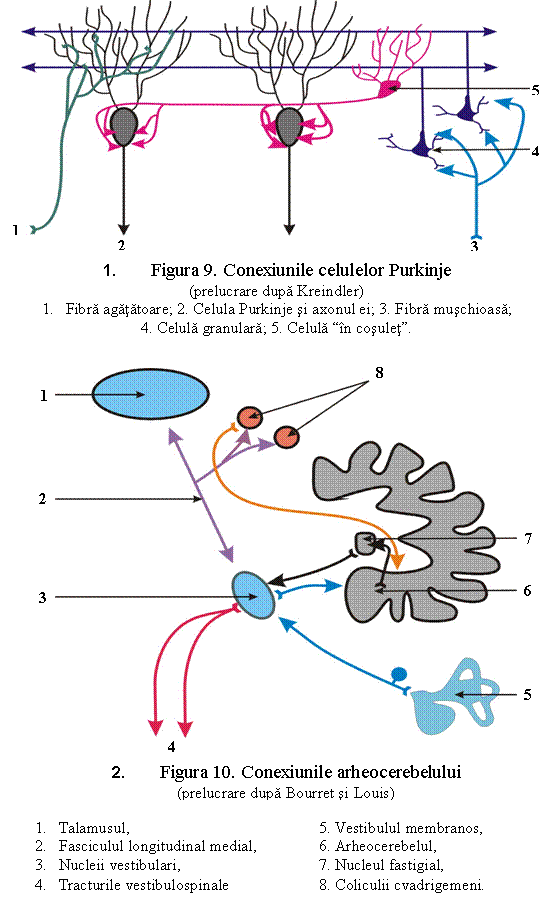
In scoarta cerebeloasa, informatiile ajung pe doua canale, adica prin doua tipuri de fibre aferente: fibre agatatoare, care patrund in scoarta cerebeloasa fara sa se ramifice, iar acolo urca pe dendritele unei singure celule Purkinje, realizand sinapse repetate cu ramurile dendritice mai mari; fibre muschioase, care, dimpotriva, se ramifica bogat in substanta alba a cerebelului, in asa fel incat aceeasi fibra da ramuri pentru numeroase lamele, ramificandu-se in cadrul unei lamele si se termina in stratul granulos, prin numeroase terminatii nervoase. Aceste terminatii "muschioase" fac sinapsa cu dendritelele scurte ale celulelor granulare (acestea formand complexe sinaptice - "glomerulii cerebelosi").
Ambele fibre aferente sunt excitatorii asupra neuronilor cu care vin in contact. In schimb, deosebirea fundamentala dintre ei consta in faptul ca o fibra agatatoare poate stimula o singura celula Purkinje, in timp ce o fibra muschioasa poate transmite excitatiile numai prin intermediul celulelor granulare. Acestea au de obicei 3-4 dendrite, deci sunt necesari stimuli simultani de la cel putin acelasi numar de fibre muschioase, ca sa fie activate. Axonul celulei granulare urca in stratul molecular, unde se ramifica sub forma literei T, apoi descrie in axul longitudinal al lamelei un traiect de 1 - 1,5 mm in ambele sensuri. Intre timp, realizeaza sinapse in stratul molecular cu dendritele celulelor Purkinje. Deci, fiecare dintre axonii celulelor granulare (fibre paralele), impreuna cu sutele de dendrite ale celulelor Purkinje care se incruciseaza cu el, formeaza "sinapse de incrucisare". Numarul celulelor granulare este asa de mare, ca pe ramificatia dendritica a unei singure celule Purkinje trec aproximativ 200 000 fibre paralele. Cum celula Purkinje are acelasi numar de dendrite, practic se formeaza sinapse cu fiecare fibra paralela. Celula granulara si prelungirile ei (fibrele paralele) sunt structuri excitatorii. Cercetatorul japonez M. Ito a descoperit ca celulele Purkinje au o actiune exclusiv inhibitorie.
Fibrele paralele stimuleaza insa nu numai celula Purkinje, ci si celula "in cosulet" din zona profunda a stratului molecular. Axonii acestor neuroni trec (sub forma de arc) perpendicular pe axul longitudinal al lamelei, jumatate intr-un sens, jumatate in celalalt sens. Astfel trimit prelungiri la fiecare din cele 10 celule Purkinje invecinate. Aceste prelungiri coborand, inconjoara originea axonului celulei Purkinje cu un plex sinaptic sub formaa de cosulet, format din terminatiile ce provin de la 30-50 celule "in cosulet". Dupa Eccles, aceasta sinapsa este inhibitorie, deci si celulele "in cosulet" sunt neuroni specifici inhibitori.
Astfel putem intelege teoria unitatilor integrative si mecanismele de conexiune ce au loc la nivelul scoratei cerebeloase. Pentru ca informatiile venite pe cale fibrelor muschioase sa ajunga in reteaua neuronala a scoastei cerebeloase, este necesar ca in acelasi timp sa soseasca o unda de excitatie pe atatea fibre muschioase invecinate cate sa puna in excitatie o celula Purkinje. In acest caz, intr-o lungime de 3 mm ajung in excitatie 300-400 de celule Purkinje. Campul minim in care celulele Purkinje ajung in asa fel in actiune este sirul de celule Purkinje situat pe o distanta de 3 mm (distanta acoperita de o fibra muschioasa). Concomitent insa, acest fascicul de fibre paralele excita si celulele "in cosulet" din calea lui, care inhiba fiecare din cele 10 celule Purkinje de pe traiectul sau. Lateral de zona celulelor Purkinje excitate exista cate un camp inhibitor pe o distanta de 10 celule Purkinje. Aceasta forma de inhibitie este denumita inhibitie colaterala. Important este ca un camp neuronal stimulat este inconjurat de un camp neuronal inhibat. Aceste campuri nu sunt, desigur, constante, ci, in functie de totalitatea informatiilor primite, se pot forma orice combinatii in oricare loc. Observatia este valabila numai pentru informatiile sosite prin fibrele muschioase.
Informatiile primite prin fibrele agatatoare ajung la celulele Purkinje aflate intr-un camp inhibat sau neutru. Unica celula pe care se termina este intotdeauna activata.
Scoarta cerebrala are ca unic element eferent celula Purkinje. Axonul ei se termina intr-unul dintre nucleii cerebelosi sau direct in nucleul vestibular lateral. Celulele Purkinje sunt elemente specifice inhibitoare; tot sistemul are sens numai daca la nucleii cerebelosi ajung cai excitatorii din alte locuri. Acestea pot veni din formatia reticulara, nucleii vestibulari sau colaterale din fibrele lungi cataratoare sau muschioase.
B. NUCLEII CEREBELULUI
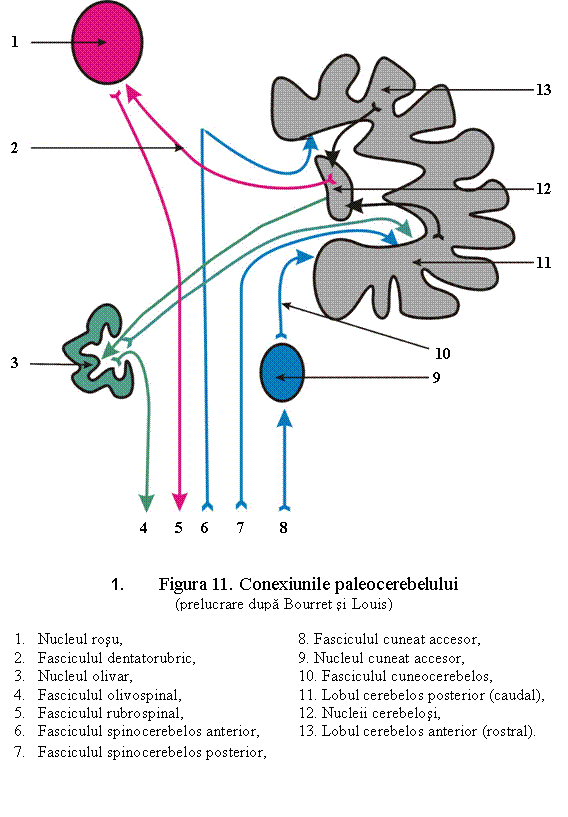
1. Nucleul fastigial (nucleus fastigii) (sin.: nucleii tavanului, nucleii cortului)
Sunt localizati la nivelul portiunii anterioare a vermisului, in zona tavanului celui de al IV -lea ventricul. Ei au forma ovoidala si contin neuroni care fac sinapsa cu axonii celulelor Purkinje (din paleocerebel) conectate la fibrele tractului cerebelos si cu fibre directe din nucleii vestibulari (tractul vestibulo-fastigial). Tractul bulbo-cerebelos, alcatuit din axonii neuronilor localizati in nucleii gracilis, cuneatus si cuneat accesor care formeaza sitemul fibrelor arciforme externe, face sinapsa in nucleii fastigiali.
Axonii neuronilor din nucleii fastigiali ajung prin pedunculul cerebelos inferior la nucleii vestibulari (fasciculul fastigio-vestibular) si formatiunea reticulara bulbara (fasciculul fastigio-vestibular), grupate sub numele de tractul fastigio-bulbar (neomologat N.I.).
2. Nucleul globos nucleus globosus) (sin.: nucleul dintat accesor intern).
Este situat lateral de nucleul fastigial. Primeste fibre din zonele paleocerebeloase.
3. Nucleul emboliform (nucleus emboliformis) (sin.: nucleul dintat accesor extern)
Este situat lateral de nucleul globos; primeste fibre de la celulele Purkinje din zonele neo- si paleocerebeloase.
4. Nucleul dintat (nucleus dentatus) (sin.: oliva cerebeloasa)
Este localizat la nivelul substantei albe a emisferelor cerebeloase. Are aspectul unei pungi cu margini plicaturate. Medial, o lama de substanta alba formeaza hilul nucleului dintat (hilus nuclei dentati). Primeste fibre predominant din cerebel dar si o mica parte din paleocerebel. Eferentele merg la nucleul rosu (tractul dento-rubric) sau direct la talamus (tractul dento-talamic).
C. CAILE CEREBELOASE
1. Caile aferente
a) Tractul spinocerebelar posterior (Flechsig) isi are originea in nucleul toracic (Clarke) din neuromerele T9-L3. Acesta primeste impulsuri de la receptorii musculari si cutanati din membrele inferioare, prin protoneuronii caii proprioceptive inconstiente. Axonii protneuronilor patrund prin radacinile posterioare ale nervilor spinali in maduva spinarii, urca 2-5 neuromere, inainte de a face sinapsa cu deutoneuronul din nucleul toracic. Calea este strict de aceeasi parte si se termina in vermis sub forma de fibre muschioase. Impulsurile provenite de la membrele superioare au o cale asemanatoare (mai putin elucidata) prin intermediul nucleului reticular lateral.
b) Tractul spinocerebelar anterior (Gowers) isi are originea in substanta cenusie laterala intermediara si la baza cornului posterior din segmentele L4-S2. Axonii lor se incruciseaza in cea mai mare parte imediat, apoi trec in partea anterioara a funiculului lateral, unde urca pana la limita dintre punte si mezencefal. Pe calea pedunculilor cerebelosi superiori, ajung in partea superioara a vermisului, terminandu-se, de asemenea, tot in fibrele muschioase.
Tractul olivocerebelar porneste din nucleul olivar din bulb, se incruciseaza pe linia mediana si, pe calea pedunculului cerebelos inferior de parte opusa, ajunge la cerebel. Dinspre toate partile olivei, dupa o proiectie stricta, fibrele ajung la o anume zona a scoartei cerebeloase. Aici se termina exclusiv prin fibre cataratoare. Prin intermediul olivei, atat caile descendente (de la talamus si nucleul rosu), cat si caile ascendente (din maduva) vin in contact indirect cu celulele Purkinje ale scoartei cerebeloase, pe baza raportului un neuron olivar / o celula Purkinje.
Tractul vestibulocerebelar este o cale incrucisata a axonilor nucleilor vestibulari destinata in special nodulusului si floculusului (arheocerebel).
Tractul pontocerebelar este o cale incrucisata importanta, alcatuita din axonii neuronilor din nucleii proprii ai puntii, care trec prin pedunculii cerebelosi mijlocii la emisferele cerebeloase si la portiunea mijlocie a vermisului (neocerebel). Caile au originea in scoarta cerebrala, de unde coboara prin tracturile frontopontin, temporopontin si occipitopontin spre nucleii proprii, care au rol de statie de releu. Defecte congenitale ale scoartei cerebrale din perioada embrionara duc la atrofia scoartei cerebeloase de parte opusa.
Scoarta cerebeloasa are un singur tip de element eferent: celula Purkinje. Axonii majoritatii celulelor Purkinje se termina in nucleii cerebelosi. Ei por cobori insa si la nucleii vestibulari laterali din bulb. Cum celulele Purkinje sunt elemente specifice inhibitorii, tot sistemul are sens numai daca in nucleii cerebelosi ajung si stimuli din alte locuri. Acestia pot veni din formatia reticulara si nucleii vestibulari, dar si cai lungi, care se termina sub forma de fibre cataratoare sau muschioase, pot da colaterale la nucleii cerebelosi.
Din nucleii cerebelosi, numeroase fibre merg la nucleii vestibulari si la formatia reticulara, dar cea mai mare parte a lor ajung prin pedunculii cerebelosi superiori in mezencefal, unde se incruciseaza si se termina in nucleul rosu. Fibrele principale se continua si se termina in nucleul ventral lateral al talamusului. De aici, un alt neuron conduce la scoarta cerebrala motorie.
CONSIDERATII ANATOMOCLINICE
Cerebelul primeste si integreaza informatii de origine periferica (senzitive si senzoriale) si centrala (corticale si subcorticale) si trimite un raspuns adecvat catre neuronii somatomotori, de obicei mediat de catre nucleii proprii ai trunchiului cerebral. Reamintim ca, din punct de vedere al conexiunilor, nucleul vestibular lateral poate fi considerat un nucleu propriu, dupa unii autori, chiar nucleu cerebelos (ANDRONESCU, 1979). Criteriul filogenetic este cel mai potrivit pentru studiul anatomoclinic al cerebelului.
1. Arhicerebelul nu primeste aferente corticale, ci prelucreaza informatiile primite de la analizatorul vestibular, urmarind repartizarea tonusului muscular in vederea mentinerii pozitiei capului in raport cu trunchiul, precum si a bulbilor oculari. Leziunile lobulului floculonodular pot produce tulburari bilaterale de echilibru static (pozitia cu picioarele departate, pentru a avea o baza larga de sustinere) si dinami: ataxia muschilor trunchiului (mers ebrios, cu tendinta de a cadea pe spate sau intr-o parte), dar fara tremuraturi si hipotonie.
2. Paleocerebelul primeste aferente somestezice bilaterale, precum si aferente auditive si vizuale prin intermediul coliculilor superiori si inferiori. Informatiile sunt integrate si se trimit semnale excitatorii sau inhibitorii asupra nucleilor somatomotori ai trunchiului cerebral. Astfel, paleocerebelul regleaza tonusul muscular postural actionand, in final, asupra muschilor axiali si extensori ai membrelor, ce se opun actiunii gravitatiei. Din punct de vedere clinic, nu se poate vorbi, la om, de un sindrom paleocerebelos. Totusi, leziunile sale pot genera o crestere a tonusului muschilor extensori, precum si reflexe posturale, asemanatoare rigiditatii prin decerebrare.
3. Neocerebelul este puternic conectat la scoarta cerebrala. Primeste aferente de la cortexul motor (pe calea incrucisata corticopontocerebeloasa) si trimite eferente in sens invers pe calea (de asemenea incrucisata) cerebelotalamocorticala la aceleasi arii ale cortexului motor de unde isi au originea caile piramidale. Desi miscarile voluntare isi au originea, in exclusivitate, la nivel cortical, cerebelul intervine in initierea, coordonarea si incetarea acestor miscari. Prin caile proprioceptive inconstiente (fasciculele spinocerebeloase anterior si posterior), cerebelul este permanent informat asupra gradului de tensiune din muschii, tendoanele si articulatiile implicate in miscarea voluntara. Astfel, el poate determina momentul cand un grup muscular poate sa intre sau sa iese din contractie, controland forta, viteza de executie, amplitudinea, directia, continuitatea miscarii. Din punct de vedere clinic, lezarea neocerebelului conduce la un tablou clinic complex, dominat de fenomenul de eliberare de sub influenta cerebelului a nucleilor ventrali si intralaminari talamici. Dimiuarea tonusului muscular (hipotonie) cu oboseala musculara marcata si nejustificata de efortul depus (astenie) pot fi insotite de tulburari si mai complexe. Membrele superioare nu pot fi mentinute constant in abductie la orizontala, deoarece informatiile de la proprioreceptori nu sunt folosite corespunzator. Pierderea coordonarii musculare (asinergia), miscari "de robot", descompunerea miscarilor, dismetria (aprecierea eronata a distantelor), adiadocochinezia (incapacitatea de a executa cursiv miscari alternative repetitive, cum ar fi pronatia-supinatia mainii), fenomenul de rebound (pierderea controlului asupra muschilor antagonisti, care determina exagerarea miscarii de apropiere a indexului de nas, cu lovirea fetei) si disartria (vorbire sacadata sau greu inteligibila) se adauga unui tablou clinic complex.
Nu trebuie sa ne mire asemanarea acestor manifestari clinice de cauza organica bine localizata cu tabloul intoxicatiei acute etanolice (starea de betie), deoarece cerebelul este deosebit de sensibil la alcool, precum si la reducerea fluxului sanguin.
|
Politica de confidentialitate |
| Copyright ©
2024 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |