HISTOLOGIA GENERALA SI HISTOGENEZA
Histologia generala si histogeneza se ocupa cu studiul organizarii si genezei tesuturilor.
Tesuturile (textus) sunt agregate de celule, sincitii, plasmodii si matrice intercelulara, organizate pentru a indeplini o functie comuna. Desi celulele sunt unitati functionale ale organismelor, in realitate, tesuturile raspund de mentinerea functionarii organismelor pluricelulare, ele reprezentand sinteza colaborarii celulelor componente individuale. Cooperarea intercelulara se efectueaza cu ajutorul unor specializari ale suprafetei celulare ca, de exemplu jonctiunile comunicans (gap).
Agregatele celulare realizeaza, in functie de aspectul general, organizare generala si insusiri functionale, patru tipuri fundamentale de tesuturi: a) tesuturi epiteliale (epitelii); b) tesuturi conjunctive; c) tesuturi musculare si d) tesutul nervos.
Histogeneza sau diferentierea tesuturilor apare in stadiul de gastrula. Incepand din acest stadiu embrionar, evolutia are un caracter calitativ, celulele formate din ectoblast, endoblast si mezoblast, generand tesuturile.
Diferentierea tesuturilor presupune anumite raporturi intercelulare si implica fenomene de adezivitate si miscari celulare, astfel incat fiecare tesut capata particularitati morfofunctionale specifice. Asamblarea tesuturilor in vederea constituirii organelor (organogeneza) incepe in stadiul de nerula.
1. TESUTURILE EPITELIALE
Tesuturile epiteliale (textus epithelialis) sunt constituite din celule strans alaturate prin intermediul unei cantitati reduse de matrice intercelulara, bogata in mucopoliglucide (sau glicozaminoglicani).
Tesuturile epiteliale separa mediul extern de cel intern si realizeaza compartimentarea mediului intern in domenii cu compozitie distincta, asigurand homeostazia mediului intern si realizarea autonomiei organismului.
Epiteliile nu sunt vascularizate (cu exceptia striei vasculare a epiteliului canalului cohlear), vasele sangvine gasindu-se intotdeauna in tesutul conjunctiv subiacent. Epiteliile sunt polarizate functional, prezentand: a) o fata asezata pe membrana bazala si b) o fata apicala, cu specializari de suprafata.
Etimologia si alcatuirea epiteliilor. In sens etimologic, termenul de "epiteliu" ( de la grecescul: epi = peste si thele = ridicatura sau mamelon) semnifica o patura celulara ce acopera o ridicatura (mamelon).
In mod conventional, membranele epiteliale se denumesc in functie de pozitia lor fata de cavitatea pe care o delimiteaza, existand: a) mezotelii, care delimiteaza cavitatile: peritoneala, pericardica, pleurala; b) endotelii, care delimiteaza lumenul cordului, vaselor sangvine si limfatice, si c) epiteliile propriu-zise, care acopera suprafetele organismului aflate in contact cu mediul extern, cuprinzand o serie de glande.
Histogeneza epiteliilor poate avea loc din toate cele trei foite embrionare.
Din ectoderm se diferentiaza: 1) epidermul si toate derivatele sale (firul de par, unghia, glandele sudoripare, glandele sebacee, parenchimul si conductele glandei mamare); 2) epiteliul senzorial al urechii interne, retina si mucoasa olfactiva; 3) glanda hipofiza (adenohipofiza); 4) medulosuprarenala; 5) epiteliul corneean si epiteliul cristalinian; 6) organul smaltului si smaltul dentar.
Din endoderm se diferentiaza: 1) epiteliul sistemului respirator; 2) epiteliul tubului digestiv (cu exceptia epiteliului cavitatii bucale si al anusului care au origine ectodermica); 3) epiteliul organelor digestive si a glandelor anexe - ficat, pancreas; 4) glandele tiroida si paratiroide; 5) epiteliul cavitatii timpanice si al tubei auditive; 6) epiteliul timusului.
Din mezoderm se diferentiaza: 1) epiteliile rinichiului si gonadelor; 2) mezoteliul; 3) endoteliul; 4) corticosuprarenala; 5) celulele epiteliale cu caracteristici speciale, ca , de exemplu, celulele Leydig din testicul, celulele luteale din ovar.
In concluzie, epiteliile pot fi definite ca: tesuturi de o mare diversitate histogenetica si structurala, dar avand drept caracter morfologic comun faptul ca sunt alcatuite din agregate stranse de celule, bogate in citokeratine, si avand ca functie fundamentala realizarea si controlarea homeostaziei mediului intern, precum si autonomia organismului fata de mediul extern.
Epiteliile se pot clasifica, avand drept criteriu structura si functia lor principala, in doua grupe de baza: 1) epitelii de acoperire si 2) epitelii glandulare. Aceasta clasificare apare oarecum arbitrara, intrucat exista epitelii de acoperire in care toate celulele secreta mucus (exemplu: epiteliul gastric) si epitelii in care celulele glandulare sunt diseminate (exemplu: epiteliul intestinal, epiteliul traheei si epiteliul oviductului). Exista, de asemenea, epitelii senzoriale, in care celulele de origine epiteliala s-au specializat in functia senzoriala (exemplu: celulele mugurilor gustativi); celulele mioepiteliale s-au specializat in functia de contractilitate (de exemplu: glandele exocrine).
1.1. EPITELIILE DE ACOPERIRE
Sunt formate din celule dispuse in straturi, care acopera suprafata organismului sau captusesc cavitatile corpului. Ele se clasifica: a) dupa numarul straturilor de celule, in epitelii simple (unistratificate), epitelii stratificate si epitelii pseudostratificate; b) dupa forma celulelor din stratul superficial (in cazul celor stratificate).
1.1.1.Epiteliile simple
Dupa forma celulelor pot fi: a) scuamoase (pavimentoase) (epithelium simplex squamosus); b) cuboidale (epithelium simplex cuboideum) si c) columnare (prismatice) (epithelium simplex columnare).
Endoteliul (endothelium), ce captuseste vasele sangvine sau limfatice, si mezoteliul (mesothelium) sunt exemple de epitelii scuamoase. Din punct de vedere embriologic si fiziopatologic, celulele endoteliale si celulele mezoteliale sunt diferite, desi au acelasi aspect la microscopul optic si reactioneaza diferit la agresiuni si in cazul proliferarii maligne.
Endoteliile formeaza o bariera activa intre sange sau limfa si mediul intestinal, realizeaza o enorma suprafata de schimb (de circa 7 300 m3 la un animal de 70 - 80 kg). De asemena, ele sunt prezente la nivelul alveolelor pulmonare si la nivelul segmentului subtire al nefronului.
Dupa forma celulelor pot fi: scuamoase sau pavimentoase, cubice si prismatice sau columnare.
Fig. 1.1. Epiteliu simplu pavimentos
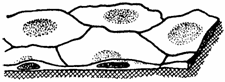
Epiteliul simplu pavimentos (scuamos) (epithelium simplex squamosus) este alcatuit din celule turtite (pavimentoase), adaptate tranzitului de gaze si lichide, cum este cazul celulelor endoteliale (endotheliocytus, i) si mezoteliale (mesothelycitus, i). In conditii patologice, cand se acumuleaza lichide in cavitati (peritoneala, pericardica si pleurala), celulele mezoteliale pavimentoase se pot transforma in celule cubice, cu platou striat (atat spre lumen, cat si spre spatiul subendotelial) si se pot desprinde, transformandu-se in macrofage. In citoplasma celulelor se pot observa numeroase vezicule de pinocitoza. De asemenea, se pot observa endotelii fenestrate prin porii transcelulari inframicroscopici si spatii intercelulare largi. Intre celulele endoteliale si membranele bazale exista un spatiu subendotelial de circa 100 Å ( = 10 nm). Ca exemple de epitelii simple pavimentoase se pot enumera: epiteliul labirintului membranos al urechii interne; epiteliul corneean posterior; epiteliul alveolar pulmonar; epiteliul foitei viscerale a capsulei Bowmann; epiteliul retelei testiculare; epiteliul spatiului subdural si epiteliul subarahnoidian (fig. 1.1.).
Fig. 1.2. Epiteliu simplu cubic
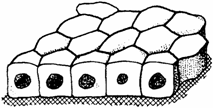
Epiteliul simplu cubic (epithelium simplex cuboideum) apare format dintr-un singur strat de celule cubice, cu nucleu sferoidal. Aceste celule isi pot modifica forma, devenind pavimentoase sau prismatice, ca in cazul epiteliului tiroidian, al glandelor mamare etc.; o forma mai constanta si-o pastreaza celulele din canalele de excretie cu lumen mic: in segmentele contorte ale tubilor uriniferi; in plexurile coroide si corpii ciliari - unde prezinta si platou striat; in bronhiolele mici (ciliate si neciliate); pe suprafata ovarului (fig. 1.2.).
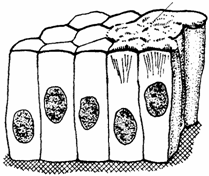
Fig. 1.3. Epiteliu simplu prismatic
Epiteliul simplu prismatic (epithelium simplex columnarum) cuprinde celule prismatice, dispuse pe o membrana bazala, intr-un singur rand. Nucleul celulelor este ovoidal, perpendicular pe membrana bazala si dispus in jumatatea bazala a celulelor. Este intalnit in cazul epiteliului mucoasei intestinale, al epiteliului ciliat al uterului si al bronhiolelor mici. Printre celulele epiteliale se observa si celule caliciforme (fig. 1.3.).
La nivelul polului apical sunt prezente complexe jonctionale ce inchid ermetic spatiul intercelular astfel incat absorbtia este posibila numai transcelular, realizandu-se o polarizare functionala foarte importanta. La nivelul polului apical se pot intalni: - platoul striat (in epiteliul intestinal, epiteliul vezicii biliare);- marginea in perie (epiteliul gastric si epiteliul tubului urinifer); - cili mobili (in epiteliul traheal, epiteliul uterin si tubar) (fig. 1.4.).
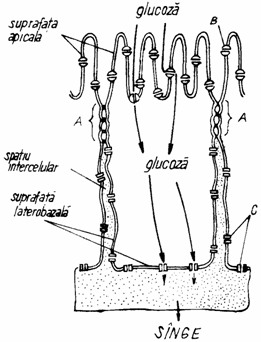
Fig. 1.4. Reprezentarea schematica a unei celule epiteliale intestinale
A - Jonctiuni stranse; B - Proteine transportoare; C - Proteine ce mediaza difuziunea facilitata.
1.1.2. Epiteliul pseudostratificat (epithelium pseudostratificatum)
Este format dintr-un singur rand de celule, care se sprijina pe membrana bazala, celule care au inaltimi diferite, astfel incat nucleii sunt plasati la inaltimi diferite. Celulele au aspecte diferite, existand celule bazale (epitheliocytus basale), intercalate (epitheliocytus intercalatus) si superficiale (epitheliocytus superficialis). Este intalnit ca epiteliu pseudostratificat prismatic ciliat in caile respiratorii si in caile genitale mascule (unde prezinta stereocili) (fig. 1.5.).
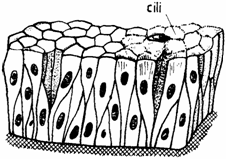
Fig. 1.5. Epiteliu pseudostratificat cilindric ciliat.
1.1.3. Epiteliile stratificate
Sunt formate din mai multe randuri de celule, celulele stratului superficial conferind denumirea epiteliului: pavimentos, cuboidal, prismatic.
Sunt epitelii de protectie pentru tesuturile subiacente sau pentru intregul organism, in cazul epiteliului cutanat. La nivelul suprafetelor umede, epiteliile stratificate pot fi: cubice, prismatice sau columnare, pavimentoase sau mixte. Suprafetele uscate sunt prevazute cu epiteliu pavimentos keratinizat sau cornificat (fig. 1.6.).
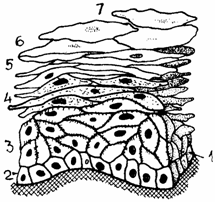
Fig. 1.6. Epiteliu stratificat pavimentos de tip cornos:
1 - Membrana bazala; 2 - Strat bazal; 3 - Strat intermediar; 4 - Strat granular; 5 - Strat lucidum; 6 - Strat cornos; 7 - Stratul celulelor descuamante.
Epiteliul stratificat pavimentos keratinizat (epithelium stratificatum squamosus cornificatum) se gaseste in epiderm, la nivelul mucoaselor bucala, faringiana si esofagiana - la rumegatoare si pasari, precum si in mucoasa prestomacelor (la rumegatoare). Este format dintr-o zona profunda, cu celule vii, si dintr-o zona cornoasa sau keratinizata, superficiala, cu celule involuate sau moarte, cu grade inalte de cornificare sau keratinizare.
Zona profunda cuprinde:
1. stratul bazal, germinativ sau generator (stratum basale) - ce contine un singur rand de celule cuboidale sau columnare (prismatice), ce se leaga de membrana bazala prin hemidesmozomi si se divid prin mitoza, dand nastere keratinocitelor care, pe masura diferentierii, trec in straturile superficiale;
2. stratul intermediar, spinos sau strat Malpighi (stratum intermedium s. spinosus), ce contine mai multe randuri de celule poliedrice, legate intre ele prin tonofibrile, ce se insera pe desmozomi, asigurand rezistenta la solicitarile mecanice.
Pe masura ce se apropie de suprafata, celulele poliedrice se turtesc si prezinta granulatii mari, bazofile, care marcheaza inceputul keratinizarii;
3. stratul granular (Unna) cuprinde mai multe randuri de celule pavimentoase, care contin granulatii bazofile de keratohialina;
4. stratul lucid - contine mai multe randuri cu aspect clar, in care granulele citoplasmatice dispar, iar nucleii intra in degenerare. Membrana celulara se ingroasa foarte mult;
5. stratul cornos sau keratinos (stratum superficiale) este format din mai multe randuri de celule cornoase sau corneocite, turtite, anucleate. Citoplasma corneocitelor este ocupata de o scleroproteina filamentoasa, birefrigerenta, denumita keratina. Corneocitele sunt solidarizate printr-un material intercelular, produs in paralel cu keratina si avand rolul de a cimenta membrana celulara. Stratul cornos este supus frecarii si pierderilor prin descuamare, pierderi ce sunt inlocuite prin diviziunile celulelor din stratul bazal.
Biogeneza epidermului
Epiteliul stratificat pavimentos de tip cornos epidermic reprezinta un prototip de epiteliu polarizat, in care pierderile prin descuamare sunt localizate la suprafata, in timp ce procesele proliferative se desfasoara exclusiv la nivelul stratului bazal. Intre cele doua straturi, keratinocitele parcurg un proces caracteristic de diferentiere, fara a se divide, comportandu-se ca o populatie de celule tranzitorii, al caror ciclu vital este limitat in timp, fiind supus unui continuu proces de reinnoire. Timpul in care keratinocitele umane parcurg procesul de diferentiere, intre stratul generator si cel descuamant, este de 2 - 4 saptamani.
In stratul bazal al epidermului se afla aproximativ 15 000 de celule pe fiecare mm². Circa 10% din aceste celule sunt celule speciale, precum: celulele pigmentare, celulele Langerhans si celulele Merkel, majoritatea celulelor fiind insa reprezentate de keratinocite, de forma prismatica (columnara), cu citoplasma bazofila, aflate in diviziune celulara. O mica parte (0,6%) din keratinocitele stratului bazal sunt celule stem, cu o mare capacitate de automentinere prin diviziune, in cursul intregii vieti a organismului, in ciuda pierderilor prin diferentiere a unei parti din populatia lor. Majoritatea celulelor din stratul bazal sunt insa celule proliferative sau precursoare, care au capacitate de diferentiere, evoluand pe linia keratinocitara. Ele provin din diviziunea celulelor stem, in care una din celulele fiice pastreaza caracterul matern, iar cea de-a doua trece in compartimentul proliferativ, angajandu-se in procesul de diferentiere. Cat timp raman in stratul bazal, keratinocitele precursoare - angajate in keratinocitare, isi pastreaza capacitatea de diviziune (fapt pentru care sunt denumite si keratinocite precursoare proliferative) - ele reprezentand principala sursa de inlocuire a keratinocitelor uzate, ritmul de diviziune a celulelor stem fiind mai lent.
Dupa cateva diviziuni in stratul bazal, care reprezinta compartimentul proliferativ, celulele precursoare trec in compartimentul functional, care incepe cu stratul spinos. Aceasta trecere este marcata de aparitia unei proteine solubile, involucrina (de la latinescul involucrum = invelis), precum si de suprimarea capacitatii de diviziune.
Compartimentul functional include straturile: spinos, granulos, lucidum si cornos, cuprinzand celulele angajate ferm in procesul diferentierii, proces care se desavarseste in stratul cornos. Celulele sunt descendentele catorva celule stem si ale unui grup (pool) relativ restrans de celule precursoare, care ocupa un sector limitat din stratul bazal.
In acest mod, biogeneza celulelor epidermice se realizeaza in unitati proliferative epidermice (UPE), reprezentate de o familie de celule, compusa din membrii cu o capacitate proliferativa si din membrii functionali. Compartimentul proliferativ al familiei este localizat in stratul bazal si cuprinde 10 - 12 celule, din care: 5 - 6 celule precursoare, una este Langerhans, iar 2 - 3 sunt celule care nu se mai divid si asteapta semnalul sa treaca in compartimentul functional. Pe fiecare mm2 de epiderm se afla circa 1 400 de unitati proliferative epidermice.
Compartimentul functional, ce include celule pe cale de diferentiere, este dispus in coloana, deasupra grupului proliferativ. Unitatile proliferative ale epidermului sunt prezente si in biogeneza epiteliilor stratificate ale esofagului, mucoasei bucale etc.
Celulele stem din stratul bazal prezinta riscul maxim de cancerizare - sub actiunea unor agenti chimici sau biologici - deoarece au un ciclu biologic semnificativ mai lent decat celelalte celule ale stratului bazal.
Procesul de keratinizare se realizeaza printr-o serie de modificari chimice, care conduc la transformarea matricei celulare in keratina, la densificarea membranei citoplasmatice si la impermeabilitatea matricei extracelulare.
Keratina provine din transformarile chimice treptate ale citokeratinelor care formeaza tonofilamentele si din keratohialina stratului granular. Citokeratina sau prekeratina se acumuleaza treptat in citoplasma celulelor stratului spinos, sub forma unor fascicule dispuse la periferia celulei. Pe masura inaintarii spre straturile superficiale, citokeratinele se sulfateaza, iar in stratul granulos, citokeratina se impregneaza cu keratohialina. Keratohialina se sintetizeaza in ribozomii liberi din celulele randurilor superficiale ale stratului spinos si atinge o mare dezvoltare in stratul granulos.
La limita dintre stratul granulos si cel cornos se formeaza α-keratina, ca rezultat al impregnarii citokeratinei cu keratohialina (fig. 1.7.).
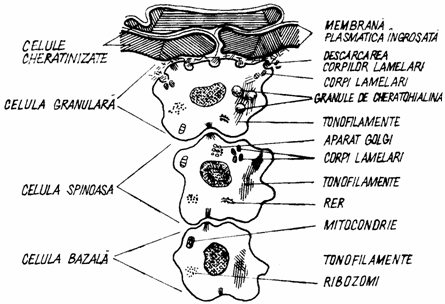
Fig. 1.7. Schema procesului de cornificare
In stratul cornos, organitele celulare sunt digerate de enzimele lizozomale. α-keratina, proteina anionica, se cupleaza cu filigrina, o proteina cationica, bogata in histidina, prezenta doar in celulele cornoase. Filigrina serveste ca schelet pentru filamentele anionice de alfakeratina. Aceasta cuplare a α-keratinei cu filigrina explica trecerea brusca de la bazofilia stratului granular la acidofilia stratului cornos. In stratul cornos, filigrina este degradata treptat in aminoacizi. Modificarea proceselor de keratogeneza are implicatii in patogeneza afectiunilor epidermice.
In fazele terminale ale diferentierii, membrana plasmatica a corneocitelor poate ajunge la o grosime de 15 nm, datorita formarii unui invelis subplasmalemal. Acest invelis devine evident la microscopul electronic, inca din stratul granular si se desavarseste in stratul cornos (fig. 1.8.).
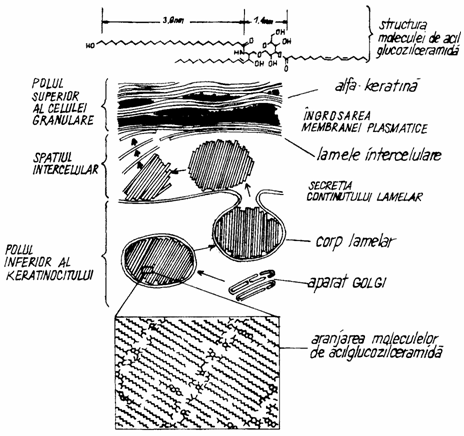
Fig. 1.8. Producerea glicolipidelor in stratul granular.
Involucrina apare ca o proteina citoplasmatica solubila, inca din stratul bazal, in keratinocitele precursoare, care isi suspenda activitatea mitotica si se angajeaza in procesul diferentierii keratinocitelor, trecand in stratul imediat superior. Involucrina solubila este un marker al constituirii epiteliilor stratificate, fiind identificata in stratul suprabazal al tuturor tipurilor de epitelii stratificate. Transformarea involucrinei in invelis subplasmalemal rezulta in urma transformarilor involucrinei solubile in involucrina insolubila si in urma complexarii (cuplarii) ei cu alta proteina si cu proteine insolubile din plasmalema sub actiunea proteinei denumite transglutaminaza.
Matricea extracelulara din stratul cornos este impermeabila si contine un material dens la fluxul de electroni, care confera o adezivitate puternica corneocitelor. Adezivitatea se datoreaza, pe de o parte, extinderii si etalarii desmozomilor, iar pe de alta parte, eliberarii in spatiile extracelulare a unui material sintetizat de keratinocite.
Keratinocitele din patura superficiala a stratului spinos, precum si cele din stratul granular, contin in citoplasma lor granule lamelare de 100 - 500 nm, denumite keratinozomi. Granulele sunt delimitate de membrane sintetizate la nivelul complexului Golgi, iar continutul granulelor este eliminat si depozitat extracelular, constituind o bariera primara pentru apa.
Epiteliul stratificat pavimentos necornificat sau de tip mucos (epithelium stratificatum squamosus noncornificatum) delimiteaza lumenul cavitatilor umede (cavitatea bucala, esofagul, corneea, cavitatea vaginala). Este lipsit de stratul cornos si cuprinde trei straturi celulare: 1) bazal (generator); 2) intermediar sau spinos; 3) superficial, in care celulele mor fara sa se keratinizeze si se exfoliaza (fig. 1.9.).
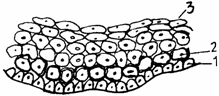
Fig. 1.9. Epiteliu stratificat pavimentos de tip moale (necornificat)
1 - strat bazal; 2 - strat intermediar; 3 - strat superficial.
Epiteliul stratificat prismatic sau columnar (epithelium stratificatum columnare) se caracterizeaza prin prezenta in stratul superficial a unor celule prismatice, inalte. Este prezent in unele zone ale conjunctivei oculare si in canalele de excretie de calibru mare ale glandelor exocrine (fig. 1.10.).
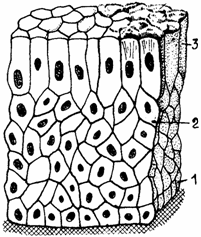
Fig. 1.10. Epiteliu stratificat columnar
1 - strat bazal; 2 - strat intermediar; 3 - strat superficial columnar.
Epiteliul stratificat polimorf (mixt, tranzitional, uroteliu) (epithelium transitionale) este caracteristic cailor urinare (ureter, vezica urinara), care sufera modificari importante de lumen si volum, consecutiv acumularii urinei. Schimbarile volumetrice impun o modificare a aspectului structural al epiteliului, prin alunecarea intre ele a celulelor epiteliale, astfel incat, in conditii de colabare, epiteliul poate prezenta mai multe straturi (10 straturi), iar atunci cand este intins, numarul straturilor se reduce (2 straturi).
In organizarea sa, uroteliul prezinta: 1) stratul bazal (generator) (stratum basale), cu un singur rand de celule cubice sau prismatice; 2) stratul intermediar (stratum intermedium) - foarte gros la rumegatoare si ecvine - cuprinde mai multe randuri de celule poliedrice si celule in forma de racheta, situate sub stratul superficial; 3) stratul superficial (stratum superficiale), format din celule mari, multinucleate, cu aspect umbeliform. In citoplasma superficiala a acestor celule se observa o densificare in crusta (bogata in fosfataza alcalina, glicogen si mucopoliglucide neutre) prin care se modifica resorbtia transepiteliala a produselor de excretie. Intre celulele epiteliale se gasesc putini desmozomi, astfel incat procesul de alunecare a celulelor se desfasoara relativ usor, permitand tranzitia rapida a celulelor dintr-un strat in altul, de unde si denumirea de tranzitional. De altfel, PETRY si AMON il descriu ca pe un epiteliu pseudostratificat cuboidal sau prismatic, in functie de gradul de distensie al organismului (fig. 1.11.).
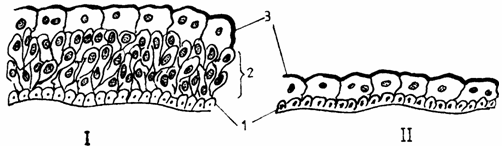
Fig. 1.11. Epiteliul de tranzitie (uroteliu)
I - in relaxare, II - in extensie.
1 - stratul bazal; 2 - stratul intermediar(celule in racheta); 3 - stratul celulelor umbeliforme.
Rolul protector al epiteliilor stratificate este conferit de structura acestor epitelii care, prin stratificarea celulelor si prin keratinizare, se opun actiunilor mecanice, impiedica evaporarea apei din organism si sunt impermeabile pentru apa si microorganisme. Acest rol protector este indeplinit si de epiteliile simple si pseudostratificate, cu ajutorul cililor si celulelor secretoare de mucus.In plus, epiteliile simple mai indeplinesc un rol: a) mecanic (epiteliul mucoasei gastrice si intestinale); b) de absorbtie (epiteliul intestinal); c) de resorbtie (epiteliul renal); d) de filtrare (endoteliile si mezoteliile); e) senzorial (celulele gustative, auditive, celulele maculelor si crestelor aparatului vestibular) si senzitiv (epiteliul olfactiv).
1.2. EPITELIILE GLANDULARE (Epithelium glandulare)
Sunt formate din celule diferentiate, celule secretorii sau glandulocite (glandulocytus) care au proprietatea de a elabora produsi specifici care parasesc celula. Impreuna cu tesutul conjunctiv, vasele sangvine si tesuturile nervoase, participa la formarea unor organe secretorii distincte, denumite glande (glandulae).
Glandele se clasifica dupa criteriul morfologic (prezenta sau absenta unui canal excretor) si functional (locul unde se varsa produsul de excretie) in mai multe grupe: a) glande exocrine (glandulae exocrinae), ce prezinta canal de excretie si isi varsa produsul la suprafata organismului sau in lumenul unor organe ce comunica cu exteriorul; b) glande endocrine (glandulae endocrinae), lipsite de canal de excretie, isi elimina produsul in mediul intern (sange, limfa); c) glande amficrine, in care aceleasi celule elaboreaza atat secretia externa cat si pe cea interna; d) glande mixte, ce cuprind grupe de celule cu secretie interna.
Dupa numarul celulelor care le compun, exista: glande exocrine unicelulare (glandulae unicellulares), reprezentate de celula caliciforma (exocrinocytus caliciformis) si glande multicelulare (glandulae multicellulares), ce pot fi endo- sau exocrine (fig. 1.12.).
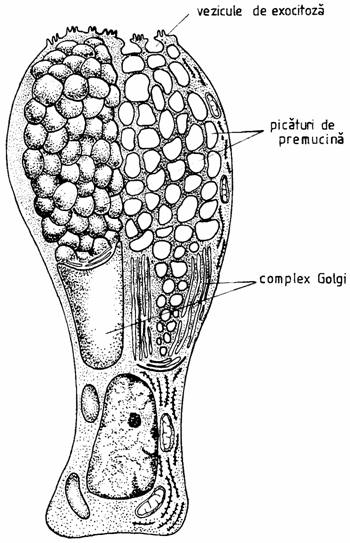
Fig. 1.12. Ultrastructura celulei caliciforme
1.2.1.Glandele exocrine
Sunt formate din unitati secretorii sau adenomere, canale excretorii, tesut conjunctiv, vase si nervi.
O unitate secretorie cuprinde un grup de celule epiteliate secretorii care delimiteaza un lumen. Canalul excretor (ductus excretorius glandulae) este un conduct epitelial care transporta produsul de secretie de la unitatea secretorie la suprafata unui epiteliu de acoperire. In unele cazuri, celulele canalului excretor pot indeplini si functii de absorbtie (fig. 1.13.).
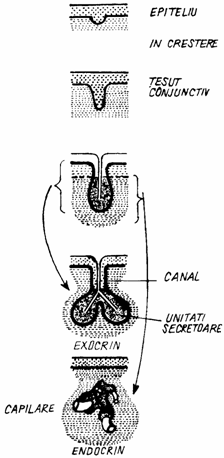
Fig. 1.13. Dezvoltarea epiteliilor glandulare
In functie de tipul canalului excretor, exista: a) glande simple (glandula simplex), care au un singur canal neramificat (exemplu: glanda sudoripara) si b) glande compuse (glandula composita), care au un sistem de canale ramificate, ca ramurile unui arbore (exemplu: glandele salivare, pancreasul exocrin).
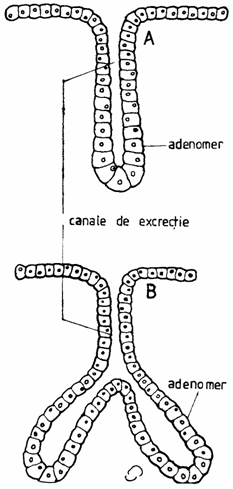
Dupa forma unitatilor excretorii, glandele exocrine pot fi: a) glande tubulare simple(glandula tubulosa) ( exemplu: glandele Lieberkühn din intestinul subtire, glanda sudoripara) sau ramificate (glandula ramosa) (exemplu: glandele Brünner din duoden) (fig. 1.14.)
Fig. 1.14. Glande tubulare
A - glanda tubulara simpla; B - glanda tubulara simpla ramificata.
; b) glande acinoase (glandula acinosa), cu unitati secretorii sferoidale (de la latinescul acinus = boaba de strugure), ca de exemplu: glandele salivare sau pancreasul; c) glande alveolare (glandula alveolaris), cand unitatile secretorii sunt sacciforme, ovoidale (in latineste alveolus = mic sac gol); d) tubularo-alveolare, cand cuprind un amestec de unitati secretorii, tubulare si alveolare (fig. 1.15.).
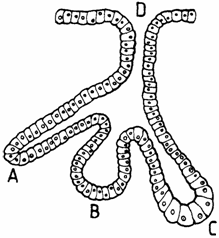
Fig. 1.15. Principalele tipuri de adenomer
A - tubular; B - alveolar; C - tubuloalveolar; D - Canal.
In functie de tipul produsului de secretie, exista urmatoarele tipuri de glande acinoase:
a) seroase (glandula serosa), care produc secretii apoase bogate in enzime; celulele seroase au baza celulei masiva, intens bazofila, datorita prezentei unui reticul endoplasmic rugos dezvoltat; nucleul celular este mare, sferoidal, iar complexul Golgi este situat deasupra nucleului, la polul apical; in citoplasma sunt prezente numeroase granule secretorii, care reprezinta vezicule de secretie, ce contin enzime si precursorii lor (fig. 1.16.);
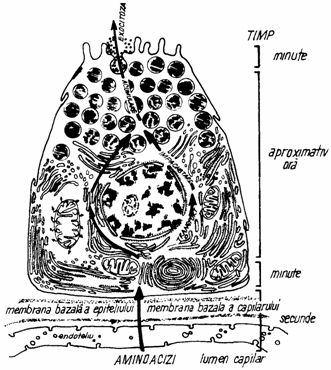
Fig. 1.16. Ultrastructura unei celule acinoase din pancreas si etapele procesului secretor.
b) mucoase (glandula mucoasa), ce cuprind celule cu nucleu mic, aplatizat, situat la baza celulei; citoplasma apare palid colorata si este ocupata de vezicule de secretii care contin mucus, ce poate fi evidentiat prin reactia PAS (Periodic Acid Schiff);
mixte sero-mucoase (glandula seromucosa), alcatuite din acini micsti, ce cuprind celule secretorii mucoase si seroase (grupate in cornuletele sau semilunele Geanuzzi (semiluna serosa) care isi varsa produsul in acelasi lumen mare, ca si al acinilor mucosi (fig. 1.17.).
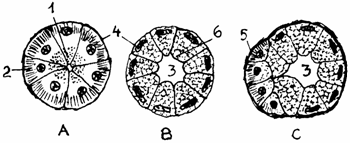
Fig. 1.17. Tipuri de acini gladulari
A - Acin seros; B. - Acin mucos; C. - Acin mixt.
1 - Granule de zimogen; 2 - Ergastoplasma; 3 - Lumen; 4 - Membrana bazala; 5 - Semiluna Gianuzzi; 6 - Mucigen.
Dupa mecanismul de secretie, glandele exocrine pot fi:
a) glande merocrine (glandula merocrina), care isi elimina produsul de secretie prin pinocitoza inversa (exocitoza) sau prin difuziune, citoplasma si membrana ramanand intacte, exemplu: glandele salivare, glandele gastrice si intestinale;
b)glande holocrine (glandulae holocrina), in care celulele participa la un singur ciclu secretor, distrugandu-se in intregime la finele acestuia, exemplu: glanda sebacee;
c) glande apocrine (glandulae apocrinae), ce pierd o parte din citoplasma apicala o data cu eliminarea secretiei; celula se reface imediat dupa excretie si reincepe un nou ciclu, exemplu: glanda mamara, singura la care apocrinia nu a fost confirmata electronooptic (fig. 1.18.).
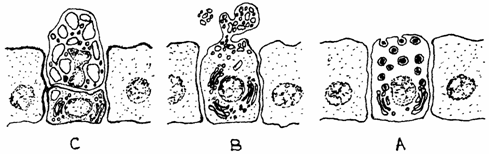
Fig. 1.18 Tipuri de eliminare a secretiei
A - glande merocrine; B - glande apocrine; C - glande holocrine.
1.2.2. Glandele endocrine
Fiind lipsite de canal de excretie, glandele endocrine isi elimina produsul de secretie in spatiul extracelular si apoi in spatiul sangvin. Celulele secretorii sunt grupate in jurul capilarelor sangvine, care formeaza o bogata retea capilara. Produc hormoni care sunt eliberati direct in sange si vehiculati spre celule, tesuturi si organe tinta.
In ultimul timp, s-a demonstrat ca si alte celule neincluse in structura glandelor endocrine au capacitatea de a secreta hormoni, ca de exemplu unele celule nervoase.
Hormonii (de la grecescul "hormaein" = a declansa o activitate, a stimula) pot avea efecte stimulatorii sau inhibatorii asupra celulelor tinta.
Exista si unele glande care elimina in sange substante folositoare , fara ca acestea sa fie hormoni, ca de exemplu: ficatul.
Din punct de vedere al dispozitiei celulare, exista glande endocrine:
- cu structura cordonala, - in care celulele sunt dispuse in cordoane intretaiate, ca de exemplu: hipofiza, suprarenala;
- cu structura veziculara, in care celulele endocrine delimiteaza vezicule sau foliculi, ca de exemplu: in tiroida;
- dispersate (sau difuze), in care celulele endocrine sunt dispersate neuniform - cum sunt glandele interstitiale din ovar sau testicule.
1.3. EPITELIILE SENZORIALE
Isi au originea in ectoderm si cuprind in compozitia lor celule dotate cu proprietatea de a culege diverse excitatii pe care le transmit cu ajutorul neuronilor la centrii nervosi corticali, unde se elaboreaza senzatiile.
Celulele epiteliale modificate si specializate pentru functia de receptie sunt de doua feluri: a) celule senzoriale propriu-zise (epitheliocytus neurosensorius), de origine nervoasa, ca de exemplu: celulele fotoreceptoare si celulele olfactive; b) celulele pseudosenzoriale (epitheliocytus sensorius), fara origine nervoasa, cum sunt celulele gustative (chemoreceptoare), celulele tangoceptoare din piele si celulele de la nivelul urechii interne.
Tesutul miopitelial este alcatuit din celule miopiteliale dotate cu proprietati contractile , fusiforme (myoepitheliocytus fusiformis) sau ramificate (myoepitheliocytus stellatus), cu miofibrile in citoplasma. Sunt prezente in glandele exocrine.
1.4. CaracterELE generale ale tesuturilor epiteliale
Tesuturile epiteliale prezinta unele caracteristici generale comune, care trec peste varietatea aspectului morfologic, a functiei si pozitiei lor in organism. Astfel, epiteliile sunt inervate de terminatii nervoase amielinice, care patrund in numar mare in epitelii si iau contact cu celulele epiteliale. Epiteliile sunt nevascularizate, cu exceptia striei vasculare din urechea interna, unde, printre celulele epiteliale, se gasesc capilare. Lipsa vascularizatiei in epitelii le obliga la o asociere tisulara cu tesutul conjunctiv subiacent, intrucat celulele epiteliale sunt aprovizionate cu substante nutritive de catre capilarele prezente in tesutul conjunctiv.
Intre epiteliu si tesutul conjunctiv se interpune membrana bazala (memebrana basalis), o structura lamelara fina a matricei extracelulare, constituita, in general, din trei straturi lamelare suprapuse: a) lamina lucida (lamina lucida), denumita si lamina rara, adiacenta celulelor epiteliale, cu o grosime de 10 nm; b) lamina densa sau lamina bazala (lamina densa s. basalis), cu o grosime de 20 - 30 nm; si c) lamina reticulata sau fibroreticulara (lamina fibroreticularis), mai putin clar definita ca strat, ce realizeaza tranzitia la tesutul conjunctiv. Membrana bazala este elaborata de celulele epiteliale, iar in structura ei s-au descris colagenul IV si V, heparan sulfatul (un glicozaminoglican) si glicoproteinele de tipul: laminina, fibronectina, entactina etc.. Proportia acestor componente variaza de la un tip de epiteliu la altul si chiar de la o regiune la alta a membranei bazale. Totusi, colagenul IV reprezinta componenta principala a membranelor bazale, fiind localizat in lamina densa.
Celulele epiteliale au o polaritate distincta, prezentand:-1) diferente intre compozitia membranei polului apical, de pe fetele laterale sau de pe fata bazala a celulei; -2) deosebiri in dispunerea spatiala a organitelor;- 3) diferentieri plasmalemale legate de functiile celulelor. Astfel, membrana polului apical a enterocitelor contine in glicocalix o proteina, ligatina, care leaga dizaharidele ce hidrolizeaza zaharurile in monozaharide absorbabile.
Polul apical al celulelor epiteliale poate prezenta diferentieri structurale care maresc suprafata de absorbttie, ca de exemplu: microvilii si stereocilii epiteliilor adaptate la functia de absorbtie (fig. 1.19.).
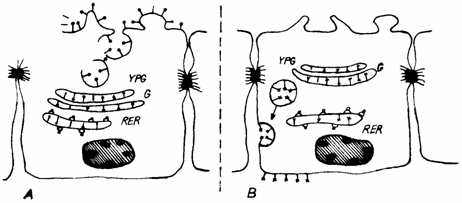
Fig. 1.19. Inglobarea glicoproteinelor la nivelul domeniului apical (A) si latero-bazal (B):
VPG - vezicule post-golgiene; G - complex Golgi; RER - reticul endoplasmic rugos(granular).
In celulele cu secretie exocrina, plasmalema apicala prezinta o permanenta activitate, participand la eliminarea secretiei printr-un proces asemanator pinocitozei inverse. La acest pol, plasmalema se remaniaza continuu. Dinamismul plasmalemei se observa si in cazul celulelor endoteliului capilarelor sangvine si limfatice, unde are loc integrarea pinozomilor cu pinocitoza inversa, determinand marirea suprafetei plasmalemale, atat inspre lumen, cat si pe suprafata dinspre tesut. In cazul celulelor hepatice si al celulelor marginale din epiteliul glandelor gastrice, plasmalema contribuie la formarea peretelui canalului intercelular sau intracelular. Si membrana latero-bazala a celulelor epiteliale contine enzime esentiale pentru absorbtia si transportul transepitelial, ca de exemplu: ATP-aza de sodiu-potasiu. (fig. 1.20.).
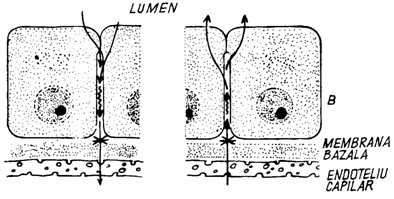
Fig. 1.20. Transportul transepitelial
A - din lumenul intestinului catre vasul sangvin; B - din vasul sangvin catre lumenul delimitat de epiteliu.
Prezenta jonctiunilor celulare distincte morfologic si functional reprezinta o alta caracteristicaa a fetelor celulei epiteliale. Ele asigura solidarizarea intima a celulelor si contureaza trecerea moleculelor prin spatiile dintre celule, realizand impartirea suprafetei celulare in sectoare distincte.
Membrana polului bazal prezinta, in cazul celulelor adaptate pentru functia de absorbtie si resorbtie (din tubul contort proximal), invaginari ale plasmalemei polului bazal, marind cu mult suprafata polului bazal (fig. 1.21.).
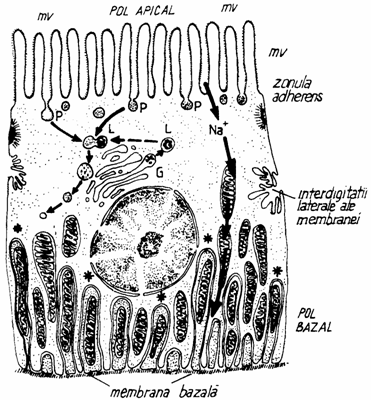
Fig. 1. 21. Schema ultrastructurii nefrocitelor din tubul contort proximal.
In celula epiteliala secretoare, membrana polului bazal contine receptori pentru hormoni, iar la epiteliile pavimentoase (scuamoase) sunt prezenti hemidesmozomii, care ancoreaza celula de matricea extracelulara.
In celulele secretorii se distinge o polaritate si in organizarea intracelulara, astfel, reticulul endoplasmic rugos (granular) este frecvent intalnit spre baza celulei, complexul Golgi este situat supranuclear, iar vacuolele de secretie apar langa polul apical al celulei. Mitocondriile se gasesc langa suprafata apicala, in celulele cu functie de resorbtie a apei si a ionilor.
Hialoplasma prezinta unele caracteristici specifice tesuturilor epiteliale. Astfel, cu toata diversitatea tipurilor de proteine de la un tip epitelial la altul, exista sase categorii de proteine intalnite in citoscheletul tuturor celulelor epiteliale: 1. actina; 2. alfa (α) si beta (β)tubulina; 3. proteine intermediare filamentare; 4. clatrina (proteina veziculelor acoperite); 5. spasmina, o proteina care se contracta in prezenta ionilor de calciu si 6. calmodulina (proteina cu afinitate pentru calciu). Filamentele intermediare sunt constituite din: vimentina, desmina, citokeratine asemanatoare alfakeratinei din epiderma. In celulele epiteliale s-au identificat 20 de tipuri de citokeratine (neutre, bazice, acide). Aceste polipeptide keratiniene sunt diferite de la un tip celular la altul, astfel ca ele difera in celulele secretorii, celulele ductale (ale conductelor) si mioepiteliale ale unei glande exocrine. Unele microfilamente intermediare sunt dispuse la periferia celulelor intermediare, ca, de exemplu, tonofilamentele - formate din citokeratine si implicate in mentinerea formei celulelor epiteliale.
In cazul tesuturilor epiteliale, metaplazia poate fi reversibila. Astfel, in anumite conditii fiziologice si patologice, un anumit tip de tesut epitelial se poate transforma in alt tip de epiteliu. Asa se intampla in hipovitaminoza A, cand epiteliile cailor respiratorii (pseudostratificate ciliate) sunt inlocuite cu epitelii stratificate pavimentoase.
Epiteliile prezinta un ciclu constant de innoire. Ele sunt structuri labile, continand populatii de celule tranzitorii ale caror celule sunt reinnoite continuu, prin activitatea mitotica. Rata de innoire este variabila, putand fi rapida (2 - 5 zile in epiteliul intestinal) sau lenta (50 de zile, in pancreas).
Studiile autoradiografice au aratat prezenta, in majoritatea epiteliilor, a unor celule stem, responsabile de producerea a noi celule. Celulele stem sunt nediferentiate si stau la baza regenerarii epiteliilor.
1.4.1. Implicarea epiteliilor in imunitate
In constitutia mucoaselor, alaturi de tesutul conjunctiv lax, se afla epiteliile, permanent expuse la contactul cu antigenii din mediul extern.
Secretiile care acopera epiteliile mucoaselor formeaza un sistem imunologic de aparare a organismului, continand in mod predominant, imunoglobulina A.
Secretia de anticorpi, la suprafata mucoaselor, este reglata independent de cea din fluidele mediului intern (plasma sangvina), lichidul pleural, lichidul peritoneal si lichidul cerebro-spinal.
Sistemul imunologic comun suprafetelor mucoase este denumit sistem imun secretor. Imunoglobulinele (A, M si E) din sucul intestinal, secretia nazala, lacrimala, sunt produse, in cea mai mare parte, din plasmocite, celule conjunctive din corionul mucoasei.
Imunoglobulinele A secretorii asigura apararea primara impotriva infectiilor locale, fapt demonstrat prin abundenta lor in saliva, lacrimi, secretie nazala, lichid prostatic. Ele nu distrug antigenii (microbi, virusi, toxine, diferite substante), ci previn patrunderea lor in mediul intern.
Imunitatea antimicrobiana are la baza diminuarea aderentei bacteriilor la epiteliile mucoaselor. Mucoasele se opun aderentei bacteriene prin secretiile lor, prin miscarea cililor (in trahee, in tuba uterina etc.), prin desmembrarea epiteliilor.
|
Politica de confidentialitate |
| Copyright ©
2024 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |
