Testiculul endocrin
1. Generalitati testicule
Testiculele sunt organe sexuale pereche, alcatuite in principal din o masa ovala de tubuli foarte spiralati acoperita de tesut conjunctiv. La fetus testiculele se dezvolta intraabdominal, la fatare sunt descinse in scrotum, dar dezvoltarea lor incepe la pubertate.
Testiculele sunt pozitionate in regiunea subanala , postpenian, cu exceptia marsupialelor, la care sunt amplasate prepenian. La majoritatea speciilor , testiculele descend in scrotum , in care raman permanent. La iepure si alte rozatoare, testiculele pot fi retrase din punga ingvinala in cavitatea abdominala chiar cand sunt dezvoltate.
Fiecare testicul este inconjurat de o capsula de fibre conjunctive colagene , tunica albuginea, din care prin extensie se formeaza de asemenea , prelungiri membranare, septule, care divid masa testiculara in lobuli alcatuiti din tubuli seminiferi.
Tubuli seminiferi prezinta o portiune puternic spiralata si o alta slab spiralata. Portiunea puternic spiralata a tubulilor seminiferi aflata langa tunica albuginea poarta denumirea de tubuli contorti. Portiunea slab spiralata a tubulilor seminiferi, aflata langa mediastin, poarta denumirea de tubuli drepti.
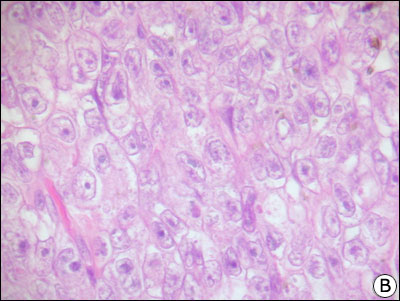
Histologic ,tubulii seminiferi sunt alcatuiti dintr-un epiteliu stratificat sprijinit pe o membrane bazala . Epiteliul este alcatuit din celule germinale si celule Sertoli. Celulele germinale se divid si se diferentiaza in spermatozoizi. Celulele Sertoli sunt mari ,cu citoplasma difuza, nedelimitata. Celulele Sertoli contribuie la formarea fluidului tubular, secreta inhibina si au rol in maturarea spermatozoizilor. Membrana bazala care inconjoara care inconjoara tubulii seminiferi contine un strat de celule contractile denumite celule mioide. Celulele interstitiale Leydig apar grupate in ciorchine printre tubulii seminiferi sau insiruite de-a lungul vaselor sanguine. La speciile cu reproductie sezoniera, celulele interstitiale se atrofiaza la sfarsitul unui sezon, diferentiindu-se altele la inceputul sezonului urmator. La unele specii sunt prezente si cellule pigmentate, care in urma atrofiei celulelor interstitiale par mai abundente si dau o culoare maro-inchis testiculelor. Celulele interstitiale reprezinta sursa primara de Testosteron.
Tubuli recti se deschid in rete testis. Rete testis se deschide in capul epididimului prin canaliculele eferente. Reteaua canaliculara din rete testis se simplfica la cateva canale care se dirijeaza spre exterior, prezentand un traiect rectiliniu intratesticular si
un traiect flexuos (ductulus eferentus).
Tubulii de rete testis sunt alcatuiti dintr-un epiteliu simplu sprijinit pe o membrana bazala. Celulele epiteliale sunt majoritatea de tip squamos dar sunt si cuboidale sau columnare in apropierea tubulilor drepti si a canaliculelor eferente.

CELULELE INTERSTITIALE LEYDIG
Testosteron
2. Sinteza de
hormoni steroizi (sexuali) 
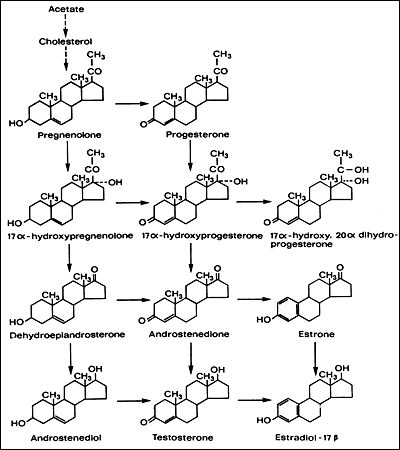
Hormonii steroizi reprezinta o clasa de hormoni care, spre deosebire de hormonii proteici, sunt lipofili. In general, ei sunt grupati in doua categorii: hormoni adrenocorticoizi ( glucocorticoizi, mineralcorticoizi) si hormoni sexuali ( estrogeni, androgeni, progesteron). Toti hormonii steroizi prezinta in molecula lor un schelet alcatuit din 17 atomi de carbon derivat de la colesterol . Desi steroizii pot fi sintetizati de novo in celula pornind de la acetat, majoritatea hormonilor steroizi sunt formati de la colesterol care este sintetizat in ficat.
Lipoproteinele cu densitate mica patrund in celula secretoare de hormoni sexuali (steroizi) in urma interactiunii cu un receptor de membrana. Sub actiunea lizozomilor, din veziculele lipoproteice este eliberat colesterolul care este utilizat imediat in sinteza de hormoni sexuali (steroizi) sau este stocat sub forma unor vezicule in celula. Primul pas in sinteza hormonilor sexuali este clivajul catenel laterale a colesterolului cu formarea pregnenolonului, ceea ce are loc in mitocondrii. Urmatoarele etape ale sintezei de hormoni sexuali se pot desfasura in mitocondrii sau pot implica miscare in alte compartimente celulare. Controlul miscarii hormonilor sexuali intre diferite compartimente celulare este inca insuficient cunoscut.
Sinteza hormonilor sexuali este conditionata de prezenta unor enzime specifice in fiecare tip de celula.
Biosinteza steroizilor sexuali presupune:
-modificarea pregnenolonului
-trecerea in stadiu de progesteron, androgeni.
Celulele care sintetizeaza androgeni (de exemplu, celulele Leydig din testicul ) poseda enzimele necesare pentru formarea pregnenolonului si a progesteronului precun si enzimele necesare transformarii progesteronului in androgeni, dar nu poseda enzimele necesare convertirii androgenului la estrogen. Celulele care sintetizeaza hormoni steroizi sexuali nu poseda enzimele pentru sinteza de hormoni corticosuprarenalieni. In schimb corticala suprarenala contine sistemele enzimatice prentru sinteza atat a steroizilor sexuali cat si a corticosteroizilor, cei din urma fiind predominanti. Aceasta face ca glanda corticosuprarenala sa produca in mod normal o cantitate mica de steroizi sexuali, cantitatea care creste considerabil in conditii patologice.
Hormonii steroizi sexuali nu sunt depozitati in celule secretoare; ei sunt eliminati imediat din celula prin difuziune transmembranara datorita propietatilor lor de liposolubilitate. Astfel , sinteza hormonilor steroizi sexuali este strans legata de excretia acestora, rata secretiei hormonale fiind controlata de rata sintezei. Singura forma de stocare a steroizilor sexuali in celula secretoare este molecula recursoare, colesterolul.
3. Transportul sangvin al hormonilor steroizi
Transportul hormonilor steroizi sexuali este mult mai complex decat al hormonilor proteici, deoarece acesti hormoni sunt lipofili, ceea ce le confera insolubilitate in solutii apoas. Acesti hormoni sunt transportati in sange pe diferite suporturi proteice. Unele proteine care leaga hormoni steroizi prezinta o afinitate mare pentru un anumit hormon steroid. De exemplu, o globulina denumita transcortina are o afinitate pentru cortizol si corticosteron dar serveste ca suport si pentru transportul progesteronului desi pentru acest hormon prezinta o afinitate mai scazuta. Proteinele de transport care prezinta o mare afinitate prezinta in schimb o slaba capacitate de transport. Albuminele plasmatice prezinta o afinitate scazuta pentru hormonii steroizi dar au o mare capacitate de transport datorita concentratiei lor mari in plasma.
Hormonul trebuie sa fie sub forma libera sau nelegat atunci cand patrunde in celula tinta pentru a isi executa rolul biologic. Aceasta impune stabilirea unui echilibru intre forma legata si forma libera a hormonului in plasma. Obisnuit, forma libera reprezinta 1% din totalul unui hormon in plasma ( o exceptie constituie cortizolul, a carui forma libera in plasma poate atinge 10% ) . Utilizarea celulara a formei libere determina disocierea hormonului legat de pe proteina transportatoare si completarea rapida a procentului de hormon liber. Obisnuit este reprezentata de cantitatea totala de hormon ( liber si legat ), exceptie facand hormonii tiroidieni iodurati. Asa dupa cum a fost aratat deja, sinteza si eliberarea hormonilor steroizi sunt strans legate. Cum rata de clerance metabolic este relativ constanta concentratia plasmatica a steroizilor reflecta fidel rata secretiei.
4. Raspunsul celular la interactiunea hormon-receptor
Hormonii steroizi sunt capabili sa interactioneze cu interiorul celulei datorita abilitatii lor de a penetra plasmalema. Interactiunea receptorului cu hormonul steroid duce la activarea complexului, care patrunde in nucleu si interactioneaza cu situsuri cromatice specifice. Rezultatul este sinteza de ARNm care , translocat pe ribozomi, directioneaza sinteza de proteine ce produc efectul biologic dorit.
5. Catabolismul hormonilor
Activitatea hormonala este limitata de catabolizarea hormonilor respectivi. Aceste procese include degenerarea sub actiunea unei varietati de mecanisme enzimatice cum ar fi hidroliza, oxidarea, hidroxilarea, metilarea, decarboxilarea, sulfatarea si glucuronizarea. Catabolizarea steroizilor implica obisnuit reducerea moleculei lor si conjugarea lor cu sulfati sau glucuronizi, ceea ce creste solubilitatea lor in apa, permitand excretarea lor prin urina. Ficatul este principalul organ responsabil de conjugarea steroizilor.
6. Spermatogeneza
Spermatogeneza incepe la pubertate si continua pana la andropauza . Spermatogeneza presupune procese de diviziune, diferentiere si reorganizare celulara care au ca finalitate obtinerea gametilor masculi ( spermatozoizii ), in care materialul genetic este redus de la starea diploida (2n cromozomi) la cea haploida (n cromozomi).
Spermatozoizii se formeaza in tubulii seminiferi, din linia epiteliala seminala, in structura careia intra urmatoarele tipuri de celule : spermatogonii, spermatocite primare, spermatocite secundare, spermatide si spermatozoizi. Fiecare tip de celula constituie stadiu evolutiv distinct al procesului de spermatogeneza. In contact cu membrana bazala a epiteliului tubulilor seminiferi se afla spermatogoniile. Celelalte celule ale liniei seminale sunt asezate in straturi succesive spre lumenul tubului seminifer, in ordinea gradului lor evolutiv. Spermatogoniile urmeaza o serie de diviziuni celulare ecuationale (mitoze) care au ca finalitate formarea spermatocitelor. Spermatocitele urmeaza diviziunea reductionala (meioza) rezultand spermatidele in care continutul in AND este redus la jumatate. Seria diviziunilor celulare de la spermatogonie la spermatida constituie spermatocitogeneza. Spermatidele urmeaza o serie de transformari structurale metamorfozice, fara diviziune celulara, la sfarsitul carora rezulta spermatozoizii. Aceste transformari metamorfozice ale spermatidelor in spermatozoizi constituie spermiogeneza.
7. Controlul spermatogenezei ( implicatiile testosteronului )
Functionarea normala a testiculelor reclama prezenta stimulatoare a gonadotropinelor a caror secretie este controlata de hipotalamus . Hipotalamusul secreta pulsatil gonadoliberin-releasing hormone (GnRH) care stimuleaza secretia hipofizara de LH si FSH din hipofiza anterioara. LH stimuleaza celulele interstitiale Leydig pentru secretia de hormoni androgeni, in principal testosteron. Androgenii determina secretia glandelor genitale anexe, stimuleaza maturarea spermatidelor si mentin singuri spermatogeneza. Principala actiune a hormonilor androgeni pare a se exercita mai mult asupra celulelor Sertoli decat asupra celulelor liniei seminale. Activitatea celulelor mioide pare a fi, de asemenea, sub controlul androgenilor.
FSH interactioneaza cu receptrorii celulelor Sertoli determinand sinteza de proteina ABP (androgen-binding-protein), conversia testosteronului la dihidrostestosteron si estrogeni, si secretia de inhibina. Proteina ABP are rol in transportul direct al testosteronului spre epididim, unde acest hormon este necesar in cantitati mari ( Hafez, 1993 ). Inhibina are efect feedback negativ asupra secretiei de FSH, nu si asupra secretiei de LH. Androgenii supreseaza secretia de GnRH, LH si FSH prin mecanism feedback negativ asupra hipotalamusului si hipofizei. Efect secundar asupra spermatogenezei exercita si alti hormoni hipofizari: PRH, GH si TSH.
8. Boli reproductive autoimune
Bolile reproductive autoimune se manifesta la debutul pubertatii cand apar noi antigene in tubul seminifer fata de care sistemul imun nu este tolerant. Toleranta fata de antigenele propii se dezvolta in timpul vietii fetale. Daca la pubertate tubii seminiferi sunt distrusi, antigenele eliberate in circulatia sistemica dezvolta un raspuns imun insotit de orhita. Orhita poate fi produsa experimental la animale prin injectarea de extracte testiculare emulsionate cu adjuvant Freund complet. In serul sangvin al acestor animale se pot identifica autoanticorpi la sperma , cand se produce distrugerea tuburilor seminiferi sau obstructia de lunga durata a canalelor seminifere. Infectia cu Brucella canis provoaca la cainii masculi o epididimita cronica. Auto-anticorpii serici sunt din clasele IgG sau IgA , care aglutineaza si imobilizeaza spermatozoizii, producand oligozoospermie sau azoospermie, urmata de sterilitate.
La vacile insamantate artificial, anticorpii spermatici s-au format in urma fagocitarii spermei de catre celulele macrofage din vagin, uter sau peritoneu, cu formarea unor cantitati sporite de auto-anticorpi ce produc sterilitate.
La unele linii de nurci negre s-a observat ca 20-30% din masculii varstnici sunt sterili, evidentiindu-se in serul sangvin nivele inalte de anticorpi antispermatici. Acesti masculi prezinta orhita cu infiltratii monocitare si depozite de complexe imune localizate de-a lungul laminei bazale a tubilor seminiferi .
9. ANDROPATII Endocrine la reproducatorii animalelor de ferma.
Frecventa din ce in ce mai mare a tulburarilor de reproductie la animalele de sex mascul a impus o dezvoltare considerabila a cercetarilor in acest domeniu. Rezultatele obtinute au fost favorizate in mare parte si de posibilitati oferite de tehnicile moderne. In stadiul actual al cunostintelor, studiul sterilitatii masculine este un domeniu vast si complex, in care sunt implicate o diversitate de factori : genetici, hormonali, metabolici etc. Daca sterilitatea masculina este relativ usor de diagnosticat, in schimb descoperirea cauzelor tulburarilor de spermato- si spermiogeneza este deosebit de dificila, deoarece este greu de desprins factorul cauzal din multitudinea influentelor nocive la care este supus organismul . De cele mai multe ori, exista un complex cauzal.
Intre componentii testiculari, epiteliul germinal este structura cea mai sensibila la numerosi factori locali sau generali, care pot afecta tranzitoriu sau definitiv procesul de spermatogeneza. In continuare, voi prezenta implicatiile endocrine in cele doua sindroame mai importante si anume: lipsa, abolirea sau pierderea libidoului si in sindromul de infertilitate prin lipsa de generare.
IMPLICATIILE ENDOCRINE IN SINDROMUL DE INFERTILITATE DETERMINATA
DE LIPSA, ABOLIREA SAU PIERDEREA LIBIDOULUI
Instinctul genetic se manifesta odata cu apritia pubertatii, fiind conditionat de o activitate hormonala complexa cu precadere a testiculelor , precum si de dezvoltarea normala a sistemului nervos. Este bine cunoscut faptul ca testiculele au pe langa functia exocrina de a elabora spermatozoizi si functia endocrina, formand androgeni in special la nivelul celulelor interstitiale Leydig. Reglarea celor doua functii testiculare se face prin circuitul de reglare hipofizo-gonadotrop. Hormonul foliculostimulator (FSH-ul) stimuleaza spermiogeneza, iar hormonul de stimulare a celulelor interstitiale (ISCH-ul) influenteaza productia androgena. La randul lor, androgenii determina comportarea sexuala, dar au o influenta si asupra epiteliului germinativ si a restului cailor si glandelor sexuale mascule. Eliberarea hormonilor gonadotropi hipofizari este reglata prin mecanisme de recuplare (feed-back negativ). Perturbari in functia testiculara (hipogonadism ) pot aparea datorita unei cedari insuficiente de gonadotropine cauzate de tulburari ale functiei hipofizare (hipogonadism, hipogonadotrop ) sau de afectiuni testiculare insotite de o productie redusa de androgeni care nu permit o franare a eliberarii gonadotropinelor hipofizare (hipogonadism, hipergonadotrop ).
Lipsa sau abolirea instinctului genetic face ca reproducatorul pus in fata unei femele in calduri sau indemnat sa efectueze saltul pe un alt mascul sau manechin, ramane indiferent sau executa saltul dupa un timp indelungat.
Tulburarile instinctului genetic constau fie in abolirea totala a instinctului, fie in scurgerea unu timp indelungat pana la efectuarea reflexelor de erectie, imbratisare si intromisiune. Ele se intalnesc mai ales la tauri si intr-o mica masura la celelalte specii.
Etiologie : lipsa sau abolirea libidoului poate fi cauzata de dereglari hormonale conditionate ereditar de catre o gena cu penetratie incompleta care face sa apara o insuficienta de ICSH.
Tratament: Utilizarea hormonilor ca tratament presupune in primul rand un raspuns al testiculului lastimulul hormonal .
Tratamentul cu testosteron ramane deocamdata medicatia de baza in cele mai multe cazuri de insuficienta sexuala. Indicatia de baza o constituie deficitul sexual cu hipoandrogenism, dar hormonul poate fi administrat cu succes si la reproducatorii cu sexualitate normala , pentru actiunea sa euforizanta toti si pentru efectul de erotizare a centrilor nervosi.
IMPLICATIILE ENDOCRINE IN SINDROMUL DE INFERTILITATE
PRIN LIPSA DE GENERARE
Impotenta de generare face imposibila monta, dar aceasta este infecunda. La baza acestui tip de infertilitate stau tulburari ale proceselor de spermiogeneza. Intre componentii testiculari, epiteliul germinal este structura cea mai sensibila la numerosi factori locali sau generali. O multitudine de factori, de noxe, pot perturba procesele de spermio- si spermatogeneza facand sa rezulte o serie de patospermii de intensitati diferite ( oligospermie , azoospermie , necrospermie ) .
Etiologie: fiind cunoscut rolul principal in controlul functiei exo- si endocrine a testiculelor al hormonilor gonadotropi ( FSH si ICSH). In cazul unei insuficiente elaborari de FSH sunt afectate procesele spermato- si spermiogeneza.
Tratament: Preparate pe baza de FSH induc primele stadii ale diviziunii celulare si mentin spermatogeneza normala ; preparatele de tip LH favorizeaza functia spermatogenetica.
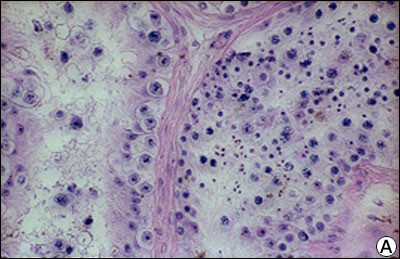
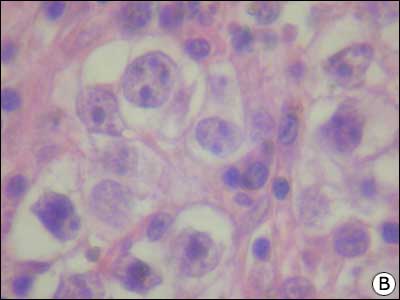
Cancer invaziv testicular - distrugerea celulelor Leydig
BIBLIOGRAFIE:
ENDOCRINOLOGIA REPRODUCTIEI LA ANIMALELE DE FERMA ; AUTOR: Boitor Ioan
FIZIOLOGIA ANIMALELOR ; AUTOR: Nicolae Dojana
TRATAT DE FIZIOPATOLOGIE GENERALA Editia a II-a ; AUTOR: Dumitru C.Curca
| Copyright ©
2024 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |
