
ACTIVITATEA ELECTRICA A MEMBRANEI CELULARE
POTENTIALUL MEMBRANAR DE REPAUS
De o parte si de alta a membranei celulare exista o diferenta de potential electric, generata de o repartitie inegala a sarcinilor electrice in cele doua compartimente. In stare stationara, in cele doua compartimente despartite de membrana se stabileste o relatie bine determinata intre toti parametrii sistemului: potential electric, potential chimic, presiune etc., ceea ce defineste echilibrul de membrana.
CONDITIILE DE ECHILIBRU
In stabilirea echilibrului de membrana intervine o serie de factori legati de caracteristicile membranei si de particularitatile mediilor disperse pe care le desparte membrana.
Organismul uman include o mare cantitate de apa (60-70% din greutatea corporala). Ea reprezinta solventul pentru sisteme disperse polifazice, cuprinzind cristaloizi sub forma ionizata (Na +, K +, Ca + + , Mg+ + etc.), molecule mici si macromolecule amfotere (in general anioni la pH fiziologic).
Mediul intracelular cuprinde si anioni proteici nedifuzibili prin membrana celulara, ceea ce impune o diferenta de potential intre cele doua fete ale membranei si deci o repartitie inegala a ionilor difuzibili
Membranele biologice au o permeabilitate diferita pentru diversele specii ionice. In aceasta situatie, ionul care difuzeaza mai usor determina aparitia unui gradient electric, gradient ce accelereaza difuzia ionului de semn opus, astfel incit, la echilibru (egalitatea potentialului electro-chimic in cele doua compartimente) nu se ajunge la egalizarea concentratiilor si anularea diferentei de potential. Echilibrul se stabileste in functie de permeabilitatea membranei pentru fiecare ion. Transportul activ impune un raport bine determinat al concentratiilor, ceea ce duce la o diferenta de potential intre cele doua fete ale membranei.
Gradientii de concentratie, de potential si de presiune existenti la un moment dat determina un transport pasiv, prin difuzie si osmoza, tinzind sa antreneze sistemul spre echilibrul termodinamic.
Transportul activ, prin procese cuplate, impune forte termodinamice constante care indeparteaza sistemul de echilibrul termodinamic, plasindu-l intr-o stare stationara.
Prin actiunea conjugata a factorilor de mai sus se ajunge la un echilibru corespunzind unei stari stationare. Asta implica:
egalitatea potentialului electro-chimic in cele doua compartimente (intra si extracelular) si deci anularea transportului net de solvit prin membrana;
izotonicitatea sistemelor disperse din cele doua compartimente si deci anularea fluxului osmotic;
neutralitatea electrica a solutiilor din cele doua compartimente.
STABILIREA POTENTIALULUI MEMBRANAR DE REPAUS
Din conditia de anulare a curentului (a transportului net) pentru toate speciile ionice, se poate deduce diferenta de potential dintre cele doua fete ale membranei, in functie de concentratie si de permeabilitate. Se obtine relatia Goldman - Hodgkin - Katz, care stabileste polarizarea membranei in repaus: potentialul de repaus (Vr):
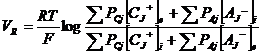
unde:
Cj = concentratia cationilor difuzibili
Aj = concentratia anionilor difuzibili
PCj = Permeabilitatea membranei pentru cationul j
PAj = Permeabilitatea membranei pentru anionul j
e si i = fata externa, respectiv interna a membranei
Principalii ioni care intervin in echilibrul membranar sunt Na +, K * si CI- si deci relatia de mai sus devine:
![]()
N.B. Prin conventie, potentialul lichidului interstitial este considerat nul si se ia ca referinta, deci potentialul fetei interne a membranei este egal cu diferenta de potential transmembranar, astfel incit sintagma 'potential membranar' e corecta si reprezinta potentialul fetei citoplasmatice a membranei.
Potentialul membranar poate fi masurat cu microelectrozi ce se introduc in celula.
Concentratia ionilor de Na+ si K+ e mentinuta de pompa de Na + /K +, compensind fluxul pasiv al respectivilor ioni.
Ionii de Cl, pentru care, in general, nu exista pompaj activ, se vor distribui in mod pasiv in cele doua compartimente, astfel incit sa fie respectat echilibrul electro-chimic (difuzia datorata gradientului de potential e compensata exact de fluxul datorat gradientului de concentratie).
Inlocuind in relatia Goldman - Hcdgkin - Katz valorile cunoscute (masurate) ale concentratiilor si permeabilitatilor principalilor ioni (Na +, K +, Cl_) se obtine, pentru diferite celule si diferite specii, un potential de - 40 -90 mV, in buna concordanta cu datele experimentale. Micile diferente sunt datorate interventiei altor ioni, existenti in concentratie mai mica (Mg++, Ca++ etc.), dar care au un rol important in realizarea functiei anumitor celule si tesuturi (exemplu: muschiul striat, muschiul cardiac).
Exemplu: in tabel sunt date concentratiile ionice aproximative pentru axonul gigant de calmar. Permeabilitatea membranei in repaus pentru principalii ioni se afla intr-un raport:
Pk : PNa: PCl = 1 : 0,04 : 0,45
|
Ionii |
Mediul citoplasmatic (mM/1) |
Mediul interstitial (mM/1) |
Potentialul de echilibru electro-chimic (mV) |
|
K + | |||
|
Na + | |||
|
Cl |
|
Concentratiile ionice tipice si potentialul de echilibru electro-chimic pentru axonul gigant de calmar
Inlocuind aceste valori in relatia Goldman - Hodgkin - Katz, la 25°C, de obtine:
VR = -61 mV
valoare foarte apropiata de datele experimentale.
CIRCUITUL ELECTRIC ECHIVALENT AL MEMBRANEI CELULARE
Membrana celulara se comporta, din punct de vedere electric, asemenea unei retele de rezistente, condensatoare si surse de tensiune electromotoare.
Concentratia principalilor ioni e diferita pe cele doua fete ale membranei celulare, diferenta mentinuta prin transport activ, ceea cc determina aparitia unei diferente de potential data, pentru fiecare specie ionica.
Gradientul de potential astfel creat determina un curent ionic pasiv, in sens invers celui activ care a stabilit gradientul, deci se comporta ca o baterie cu tensiunea electromotoare (t.e.m.) egala cu potentialul Nernst.
Pe de alta parte, membrana opune o anumita rezistenta trecerii ionului: acesta pierde o parte din energia lui datorita ciocnirilor cu peretii canlului. Se poate deci defini o rezistenta R1, a canalului si o conductanta g1 = 1/R1. Conductanta totala a unitatii de suprafata a membranei e data de produsul dintre conductanta unui singur canal si densitatea canalelor deachise (N) pentru fiecare specie ionica
g = g1N
Cu alte cuvinte, conductanta membranei defineste in termeni electrici permeabilitatea ei.
Fluxul ionilor prin membrana este astfel determinat de combinatia in serie a unei surse de t.e.m. a carei valoare e data de potentialul de echilibru electro-chimic, deci dependenta de raportul concentratiilor, si o rezistenta, expresie a permeabilitatii membranei.
Consideratiile de mai sus sunt valabile pentru fiecare specie ionica in parte, deci schema electrica echivalenta a unei portiuni de membrana va cuprinde cite o ramura baterie -rezistenta corespunzatoare fiecarui tip de ion, legate in paralel.
Pe suprafata membranei, asa cum am
vazut la difuzia particulelor incarcate, se acumuleaza sarcini electrice. Ele
sunt despartite de un mediu dielectric, format de stratul bilipidic, a carui
suprafata e de circa 100 de ori mai mare decit a tuturor canalelor la un loc.
Acest lucru confera membranei si o comportare capacitiva, deci schema electrica
echivalenta trebuie completata cu un condensator (C). In consecinta, pe linga
fluxurile ionice, prin membrana apare si un curent capacitiv de
incarcare/descarcare a condensatorului, curent dat de deplasarea de sarcini (Q)
la suprafata membranei, proportional cu valoarea capacitatii (C) si cu viteza
de variatie a potentialului (V):
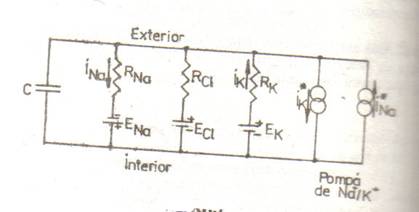
N.B. Schema electrica echivalenta descrisa se refera, in mod expres, la celula neuronala si la cea musculara striata. Membrana plasmatica a altor celule excitabile poate prezenta diferente semnificative, constind in: implicarea si a altor ioni (exemplu: calciu); interventia altor pompe (exemplu: pompa de schimb Na+/Ca+ +); valori diferite ale parametrilor etc.
EXCITABILITATEA MEMBRANEI CELULARE
Una din principalele caracteristici ale organismelor vii estc reactivitatea, deci capacitatea de a raspunde variatiilor mediului ambiant.
Excitantul (stimulul) reprezinta o variatie a proprietatilor fizico-chimice ale mediului, care poate determina o modificare fiziologica a parametrilor sistemului biologic. Pentru a fi eficient, stimulul trebuie sa fie adecvat sistemului respectiv si sa aiba anumite caracteristici de intensitate, durata, bruschete, etc. In cercetarile experimentale excitantul cel mai utilizat este cel electric, deoarece e adecvat majoritatii sistemelor biologice si poate fi dozat si masurat cu precizie.
Una din structurile excitabile este membrana plasmatica a anumitor celule, cum ar fi neuronul, fibra musculara. celulele senzoriale,etc.
Excitatia imbraca aspecte diferite:
electrice: modificarea parametrilor electrici ai membranei celulare (impedanta, polarizare, curenti transmembranari);
optice: modificari de transparenta, indice de refractie, polarizare a luminii;
radiante: emisie de radiatii infrarosii si uneori vizibile si ultraviolete;
chimice: reactii biochimice:
termice: producerea de caldura.
MANIFESTAREA ELECTRICA A EXITATIEI CELULARE. POTENTIALUL DE ACTIUNE
Aspectul electric al excitatiei celulare consta in variatia tranzitorie a potentialului membranar ca raspuns la stimul, prin modificarea permeabilitatii membranei pentru diferite specii ionice si antrenind curenti transmembranari. Cel mat adesea (celulele neuronale, musculare), raspunsul consta in depolarizarea celulei, deci micsorarea electronegativitatii fetei interne a membranei si chiar pozitivarea ei fata de lichidul interstitial. In cazuri speciale apare a depolarizare spontana a membranei. La alte tipuri de celule, membrana se hiperpolarizeaza ca raspuns la stimul.
Exista doua tipuri de raspuns electric al membranei la actiunea unui excitant:
potential electrotonic (local):
potential de actiune (PA).
Mecanismul de generare a raspunsului electric membranar. Caracteristici generale
In repaus, membrana celulara e foarte putin permeabila pentru ionii de Na+, mentinuti in concentratie mare in exterior, prin transport activ; in schimb ea este permeabila pentru ionii de K+, care ies pasiv din celula.
Aparitia unui stimul determina o depolarizare locala a membranei. ceea ce duce la deschiderea canalelor de sodiu (comandate electric) si, in consecinta, la patrunderea ionilor de Na in celula in sensul gradientului electro-chimic. Urmare a influxului de sarcini pozitive, membrana se depolarizeaza mai mult. Efluxul ionilor de K+ si transportul activ introduc o reactie negativa, avind tendinta de a stabili echilibrul.
S-a constatat experimental ca exista canale distincte pentru Na + si K +, cu o selectivitate foarte mare (principiul independentei fluxurilor ionice). Ambele sunt activate (deschise) si dezactivate (blocate) de constituent polari, sensibili la modificarea potentialului membranar si care determina deplasari de sarcina in membrana, permkind or' impiedicind trecerea ionilor. Deschiderea canalelor de potasiu e mult mai lenta decit a canalelor de sodiu. In plus conductivitatea pentru K+ (gK) nu mai poate creste prea mult, ea fiind mare si in repaus.
Exista o intirziere a declansarii raspunsului fata de momentul aplicarii stimulului, intirziere numita perioada de latenta. Ea depinde de constantele de timp ale reactiilor biochimice care duc la deschiderea canalelor dc sodiu, de caracteristicile stimulului si de mecanismele membranare implicate.
Potentialul electrotonic
Potentialul electrotonic (local) reprezinta raspunsul la stimuli slabi, aflati sub un anumit prag (stimuli subliminari), si caracterizeaza in special zone specializate ale sistemului nervos: membrana postsinaptica - potentialul postsinaptic excitator sau inhibitor si celulele senzoriale, unde formeaza potentialul de receptor si/sau generator Se manifesta prin depolarizarea membranei ori, in anumite cazuri, hiperpolarizarea ei. Este un semnal analogic, modulat in ampiitudine: creste cu intensitatea stimulului. Depolarizarea se propaga pe distante mici (maximum mm), cu pierderi (propagare decrementala). Dupa o depolarizare usoara, potentialul revine la valoarea initiala de echilibru, datorita reactiei negative date de efluxul de K+.
Potentialul de actiune
Este caracterisitic sistemului nervos central (SNC). fiind manifestarea electrica a influxului nervos. Are urmatoarele caracteristici principale:
Apare atunci cind intensitatea excitantului depaseste un anumit prag (sitmul supraliminar).
Reprezinta o depolarizare puternica a membranei, pina la pozitivarea fetei interne.
Are caracteristici strict determinate (amplitudine, forma etc.), indiferent de intensitatea excitantului (cu singura conditie de a depasi pragul): PA este deci de tipul 'tot-sau- nimic', in sensul ca ori nu apare deloc ori apare avind aceleasi caracteristici.
Intensitatea stimulului se traduce, in general, in frecventa de repetitie a PA; este deci un semnal discret. modulat in frecventa.
Se propaga in toate directiile, la distante mari, fara pierderi (de fapt nu e o propagare a semnalului electric, ci o propagare a excitatiei).
Valoarea pragului difera de la o celula la alta. Mecanismul generarii PA si, in consecinta, forma lui sunt deosebit de complexe, recunoscind mai multe faze
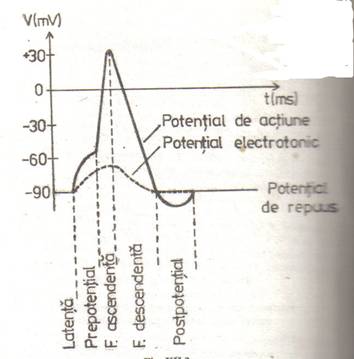
Prepotential. La inceput, depolarizarea creste lent, ca si in cazul potentialului local.
Faza ascendenta. La atingerea unei depolarizari critice se declanseaza un proces in avalansa (proces cu reactie pozitiva): patrunderea sodiului in celula determina depolarizarea membranei celulare, ceea ce, la rindul ei, duce la o crestere a permeabilitatii (deschiderea unui numar mare de canale) si deci la un influx mai masiv de Na+; depolarizarea se accentueaza, permeabilitatea creste din nou s.a.m.d.
Pragul la care se declanseaza potentialul de actiune corespunde depolarizarii la care influxul de Na+ este egalat exact de efluxul de K+. Este deci o stare labila, de la care sistemul poate evolua in doua directii: efluxul de K+ depaseste cu putin influxul de Na +, sistemul revine prin reactie negativa, la starea stationara de repaus; in caz contrar, deci la o
crestere a curentului de sodiu, apare o reactie pozitiva. data de influxul masiv de Na , in sensul gradientului electro-chimic deosebit de puternic gradientul de concentratie si de potential au acelasi sens).
Transportul activ, mai lent, si efluxul de K nu mai pot compensa influx de Na+ atit de rapid. Rezultatul e o depolarizare, puternica a membranei, pozitivarea ei pina aproape de potentialul de echilibru electro-chimic al sodiului, care insa nu e atins.
Faza descendenta. Spre sfirsitul fazei ascendente. scade din nou permeabilitatea membranei pentru Na, prin blocarea specifica a canalelor datorita noii valori a potentialului membranar. Curentul de K devine preponderent, cu atit mai mult cu cit creste si permeabilitatea membranei pentru potasiu, care iese masiv din celula, contracarind influxul de sarcini pozitive. Aceste mecanisme determina scaderea. mai lenta, a depolarizarii pina la potentialul de repaus si. uneori. inducind chiar o usoara hiperpolarizare, numita postpotential, datorita efluxului mai puiernic de K.
Concentratiile intra si extracelulare de sodiu si de potasiu vor fi stabilite prin transport activ (pompa dc Na - K ).
Perioada refractara. In faza ascendenta si in cea mai mare parte a fazei descendente a PA, membrana e insensibila la aplicarea unui nou stimul: perioada refractara absoluta. In faza ascendenta, procesul in avalansa fiind oricum declansat, un al doilea stimul nu mai poate avea nici un efect.
Spre sfirsitul fazei descendente insa si in timpul postpotentialului, membrana prezinta o excitabilitate scazuta - perioada refractara relativa - dind un raspuns de amplitudine mai mica.
N.B. Consumul eel mai mare de energie are loc la restabilirea concentratiilor de repaus, deoarece aici intervine transportul activ, celelalte faze sunt rezultatul unui complex joc al permeabilitatii, prin deschiderea si inchiderea canalelor, implicind deci transport pasiv.
Raspuns repetitiv. Daca intensitatea stimulului este mare sau excitatia e de lunga durata, membrana neuronala raspunde printr-o succesiune de PA. Frecventa de repetitie creste cu logaritmul intensitatii si e limitata de perioada refractara.
Se vede asadar ca procesul de declansare a PA reprezinta a conversie analog-discreta, semnalul analogic modulat in amplitudine (stimul sau potential electrotonic) e convertit intr-un semnal discret modulat in frecventa (succesiunea PA, impulsuri avind caracteristici identice). Transmiterea informatiei prin impulsuri modulate in frecventa e caracteristica SNC.
Acomodarea reprezinta fenomenul de crestere a pragului de excitabilitate a fibrei nervoase, atunci cind asupra ei actioneaza un stimul de lunga durata.
PROPAGAREA EXCITATIEI. MECANISM
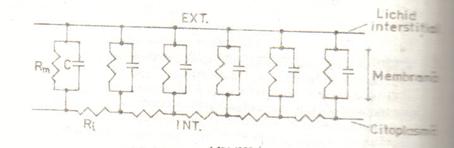
Propagarea pasiva. Proprietatile de cablu ale membranei
Pentru studiul propagarii excitatiei de-a lungul membranei axonale. trebuie avuta in vedere schema electrica echivalenta. O asemenea schema se obtine prin inlantuirea circuitelor echivalente ale fiecarei portiuni de membrana, legate intre ele prin rezistente Ri ale citcplasmei. Rezistenta lichidului interstitial e neglijabila, dat fiind volumul mare de electrolit. Se obtine astfel o structura de cablu cu elemente distribuite, reprezentata simplificat in Fig. S-au neglijat sursele de t e.m , iar rezistentele canalelor au fost inlocuite prin rezistenta lor echivalenta:
![]()
Propagarea regenerativa. Curentii locali
Depolarizarea membranei celulare intr-un punct (acolo unde se aplica sitmulul) duce la aparitia unei diferente de potential intre punctul respectiv si zonele invecinate. Ca urmare, se va produce o deplasare a sarcinilor pozitive spre regiunile mai electronegative si deci depolarizarea lor. Daca depolarizarea atinge valoarea prag, se va genera un nou potential de actiune. Se inchid astfel circuite locale de curent, numite curenti locali sau curenti Hermann.
Viteza de propagare
Cu cit distanta pina la care stimulul reprezentat de curentii Hermann este mai mare, cu atit viteza de propagare a excitatiei creste.
Mielinizarea fibrelor nervoase determina o rezistenta mare a membranei Rm. Viteza de propagare saltatorie, de la un nod Ranvier la altul. ajunge la 100 m/s. Rezulta astfel ca la animalele superioare se obtine o viteza de propagare apreciabila, fara marirea dimensiunilor fibrei nervoase.
|
Politica de confidentialitate |
| Copyright ©
2024 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| SISTEMUL COMPLEMENT |
| Metabolismul aminoacizilor |
| Cromozomul uman |
| ORGANIZAREA MOLECULARA, BIOCHIMICA A CELULEI |
| SISTEMUL MUSCULAR |
| CAILE RESPIRATORII |
| Definiția lipidelor membranare |
| MORFOLOGIE BACTERIANA |
| Termeni si conditii |
| Contact |
| Creeaza si tu |