
MEMBRANA CELULARA - ACTIVITAȚI FIZIOLOGICE ȘI IMPORTANTA IN TRANSPORTUL IONILOR ȘI AL MOLECULELOR MICI
Transportul membranar
1. Transport pasiv - conform gradientilor (concentratie, potential electric)
Difuzie simpla
Difuzie simpla prin membrana → substante hidrofobe
Canale ionice → ioni
Difuzie facilitata
Transportori → ioni, molecule mari (ex: glucide)
2. Transport activ - in sens contrar gradientilor; necesita consum de energie
Transport activ primar
Pompe ionice → ioni
Transport activ secundar
simport → ioni, molecule mari
antiport → ioni, molecule mari
3. Macrotransport
Trafic vezicular → proteine, alte molecule mari
Transportul prin membrana celulara poate fi de 2 tipuri:
Transportul ionilor și moleculelor mici care sunt determinate de proprietațile de permeabilitate ale membranei (transportul prin membrane).
Transportul macromoleculelor și proteinelor care se face prin vezicule de endocitoza și exocitoza (transportul cu membrane).
Transportul prin membrane se face prin intermediul proteinelor de transport membranar care sunt: proteine carrier- care leaga specific o molecula suferind apoi o modificare conformaționala in asa fel incat vor elibera molecula pe fața opusa a membranei.
Proteine canal care formeaza pori hidrofili ce traverseaza bistratul lipidic, cand acesti pori sunt deschisi, ei permit trecerea unor molecule specifice (de obicei ioni anorganici) de pe o parte in cealalta a membranei.
Toate proteinele canal si o parte din cele carrier permit trecerea substantelor in mod pasiv, adica in sensul gradientelor de concentrație sau electrice (gradient electrochimic) procesul se numeste transport pasiv (difuzie simplu sau difuziune facilitata).
Membranele celulare mai conțin și proteine capabile sa pompeze moleculele contra gradientului electrochimic, fiind denumit transport activ, acesta este mediat numai de proteine de tip " carrier".
Transportul activ este posibil pentru ca este cuplat cu o sursa de energie metabolica.
Difuziunea simpla a moleculelor prin membrane
In difuziune simpla o molecula aflata in soluție apoasa prin membrana, rata de difuziune este direct proportionala cu gradientul de concentrație. Moleculele mici hidrofobe trec rapid prin membrana, iar moleculele incarcate electric indiferent de marimea lor nu pot traversa membrana prin difuziune.
Transportul mediat de proteine carrier
Are la baza legarea moleculei ce urmeaza a fi transportata de o proteina transmembranara. O proteina de tip carrier are un situs specific de legare a substantei de transport cu rol de substrat. Daca apar inhibitori competitivi care se leaga specific de situsurile active atunci transportul poate fi blocat.
Sistemele de transport pot fi:
sistem uniport- transporta un singur tip de molecula de pe o fața pe alta a membranei.
Sistemul de transport cuplat cant transportul unei molecule depinde de transportul simultan al alteia. Daca intra in acelasi sens = simport, daca se face in directii opuse = antiport.
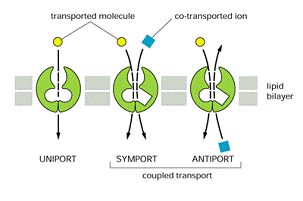
Exista o serie de substante care pot fi transportate fie pasiv, fie activ functie de concentratie si de importanta lor la nivel celular. Dar spre deosebire de difuziunea pasiva, in difuziunea facilitata pasajul moleculelor care se dizolva in bistratul lipidic este mediat de proteine ce transporta moleculele prin membrana, fara sa interacționeze direct cu interiorul hidrofob. Prin acest mecanism se transporta molecule polare și cu sarcina electrica cum sunt monozaharidele, aminoacizii, nucleotidele și ionii.
Difuziunea facilitata este mediata de doua clase de proteine distincte:
Proteine carrier
Proteinele carrier se pare ca sunt proteine transmembranare care iși modifica conformația in mod reversibil, schimbarea conformației fiind generata de legarea de molecula de transportat.
Exp : transportul glucozei/ transportul unor ioni (banda 3 din structura membranei eritrocitare funcționeaza ca un sistem antiport pentru ionii de Cl- și HCO3).
Proteine canal -formeaza porii membranari care permit trecerea oricarei molecule de dimensiune și sarcina adecvata.
Transpotul glucozei prin difuziune facilitata
Transportul glucozei (sursa principala de energie metabolica) este asigurata de o proteina carrier bine studiata
Difuzie facilitata (transport pasiv)
Transport activ glucoza, dupa principiul simport, impreuna cu Na+,
Astfel, glucoza este preluata din spațiile extracelulare, unde concentratia este mare, de catre celulele animale, prin transport pasiv cu ajutorul proteinelor carrier. Acest transport este uniport si se numeste difuziune facilitata.
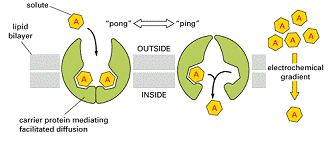
Celulele renale și cele intestinale trebuie sa preia glucoza din lumenul intestinal si respectiv tubii uriniferi, unde concentrația este scazuta. Aceste celule transporta activ glucoza, dupa principiul simport, impreuna cu Na+, a carui concentrație extracelulara este foarte mare.
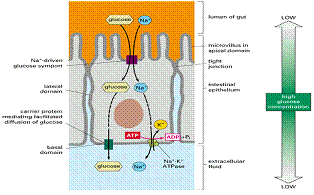
In eritrocite transportorul de glucoza este o proteina de 55kd, care funcționeaza alternand 2 stari funcționale. In starea A situsul de legare este orientat spre exteriorul celulei , legarea glucozei de acest situs, induce schimbarea conformației transportorului, care trece in starea B și elibereaza glucoza in celula.
Aceasta modificare a conformației transportorului are loc in celulele hepatice dupa un pranz bogat. Cand nivelul glucozei din sange scade, glucagonul stimuleaza hepatocitele sa produca mari cantitați de glucoza prin glicogenoliza, ca urmare concentrația glucozei din celule este mai mare decat in exterior și are loc transportul.
Canale ionice
Formeaza pori deschisi prin care moleculele cu dimensiuni și sarcina adecvate pot trece liber prin bistratul lipidic.
Exp:- Porinele din membranele bacteriilor
Joncțiunile gap - sunt proteine care delimiteaza canalele prin care sunt transportate molecule intre doua celule vecine.
Aquaporine - proteine canal pentru apa
Canalele ionice sunt cele mai bine studiate din categoria proteinelor canal, in special in celule nervoase și musculare unde deschiderea și inchiderea lor asigura transmiterea semnalelor electrice.
Prezinta 3 caracteristici funcționale:
Transportul prin canale este extrem de rapid- printr-un canal deschis trec mai mult de 1 milion de ioni/sec. (de 1000 x mai mulți decat prin prot. Carrier)
Permit un transport selectiv
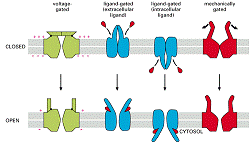
Majoritatea canalelor nu sunt deschise permanent (unele se deschid ca raspuns la legarea neurotransmitatorilor= canale cu poarta dependenta de ligand, iar altele se deschid ca raspuns la schimbarea potențialului transmembranar= canale cu poarta dependenta de voltaj.
Pentru ca funcționeaza tranzitoriu, la comanda modificarilor de membrana se numesc canale cu poarta. Au fost descrise pana in prezent 50 de canale cu poarta care sunt responsabile in special de excitabilitatea electrica nervoase și musculare și mediaza toate formele de semnalizare electrica din sistemul nervos.
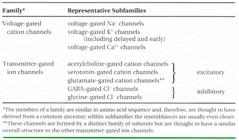
Tipuri de canale cu poarta:
Comandate de voltaj (deschiderea e cauzata de modificarea potențialului de membrana)- canale de K+, de Na+ și Ca2+
Comandate de un stimul mecanic de la suprafața celulei (canale de K+= din membranele celulelor senzoriale din urechea interna a vertebratelor).
Comandate de un ligant sau molecula semnal care este mediator extracelular (un neurotransmițator, receptorul pentru acetilcolina) sau un mediator intracelular (ioni -Ca2+; nucleotide (GMP-ciclic); proteine reglatoare (G).
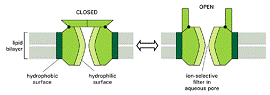
Schema generala unui canal ionic
Cai de funcționare a canalelor ionice
Canal comandat de voltaj


Toate celulele conțin pompe ionice ce folosesc energia obținuta prin hidroliza ATP pentru a transporta activ ioni prin membrana plasmatica. De aceea compoziția ionica a citoplasmei este diferita de cea a lichidelor extracelulare.
De exemplu, Na+ este pompat activ din celule, pe cand K+ este pompat activ in celule. Deoarece ionii sunt incarcați electric, transportul lor prin membrana conduce la stabilirea unui gradient electric transmembranar.
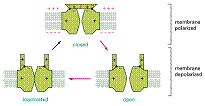
In repaus membrana este mai permeabila pentru K+ decat pentru Na+ sau pentru alți ioni, fluxul de K+ avand cea mai mare contribuție la stabilirea potențialului membranar de repaus.
Pe masura ce impulsurile nervoase (potențialul de acțiune) se propaga de-a lungul axonului, membrana se depolarizeaza. Potențialul de membrana creste de la -60mv la +30 mv, in mai puțin de o milisecunda, dupa care devine din nou negativ, revenind la valoare de repaus.
Aceste schimbari rezulta din deschiderea rapida și secvențiala a canalelor de Na și K, dependente de voltaj. Intrarea brusca a Na+ duce la o creștere rapida a potențialului de membrana pana la +30mV (apropiindu-se de potențialul de echilibru al Na+= +50mV). In acest moment se inactiveaza canalele de Na+ și se deschid cele de K+, crescand permeabilitatea pentru K+
Depolarizarea regiunilor adiacente ale membranei plasmatice permite conducerea potențialului de acțiune de-a lungul axonului neuronului ca semnale electrice, impulsurile nervoase fiind transmise rapid la distanțe mari (de peste 1m, la motoneuronii umani).
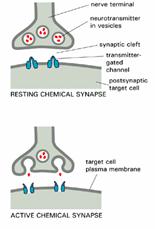
Sosirea potențialului de acțiune la nivelul butonului terminal determina deschiderea canalelor de Ca2+ dependente de voltaj, cresterea concentrației Ca2+ și eliberarea neurotransmițatorului (acetilcolina), care transporta semnalele intre celule și sinapse. Astfel semnalul electric este transformat in semnal chimic.
Acetilcolina se leaga de receptorii de pe membranele postsinaptice și deschide canalele pentru Na+ și K+dependente de ligant.
Intrarea rapida a sodiului depolarizeaza membrana celulei musculare, declanșand potențialul de acțiune, care deschide canalele de Ca2+, dependente de voltaj din reticulul endoplasmatic determinand contracția. Deci semnalul chimic este reconvertit in semnal electric.
Transportul activ cuplat cu hidroliza ATP
In transportul activ energia furnizata de hidroliza ATP este folosita pentru a dirija transportul moleculelor impotriva gradientului electrochimic, intr-o direcție nefavorabila energetic.
O mare parte din energia celulara este folosita pentru menținerea gradientelor de concentrație a unor ioni cum ar fi Na+, K+, H+, Ca2+.
Au fost descrise trei clase principale de enzime care cupleaza hidroliza ATP cu transportul ionilor impotriva/ in sensul gradientului electrochimic (vezi tabel).
|
Enzime care cupleaza hidroliza sau sinteza ATP cu transportul ionilor impotriva/in sensul gradientului electrochimic |
Tipul transportului |
|
ATP-azele din clasa P = polipeptide transmembranare care se fosforileaza in timpul transportului |
Na+, K+-ATPaza Ca2+ -ATPaza ATP-aze transportoare de protoni |
|
ATP-azele din clasa V =monomeri polipeptidici care desi hidrolizeaza ATP -ul nu se fosforileaza in timpul transportului |
Transportori de protoni (H+) localizati in membrana lizozomilor, menținand ph scazut in interiorul acestora. Transportul se face contra gradientelor de concentrație (activ) |
|
ATP-azele din clasa F= ATP-sintetaze Se afla in membrana mitocondriilor |
Transporta protoni in sensul gradientului de concentrație, transport cuplat cu sinteza ATP din ADP și fosfat |
Citoplasma are o concentrație mare de molecule osmotice cum ar fi glucoza, aminoacizii și nucleotidele. Din aceasta cauza in lipsa unui echilibru osmotic, apa tinde sa intre in celula prin osmoza ceea ce ar provoca umflarea și chiar spargerea celulei.
Echilibrul osmotic necesar este furnizat de gradiente de ioni, stabilite de catre pompa Na+/K+. Astfel pompa stabilește o concentrație mare de Na+ in afara celulei. Iar pasajul K+ prin canalele deschise stabilește un potențial electric transmembranar.
Pentru fiecare molecula de ATP hidrolizata, se transporta spre exterior 3 ioni de sodiu și spre interior 2 ioni de potasiu.
Proteina care formeaza pompa de sodiu si potasiu funcționeaza dupa sistemul antiport, ea pompeaza ionii ca urmare a unor schimbari conformaționale ciclice. Aceste schimbari sunt analoage cu cele de la difuziunea facilitata, dar sunt induse de un ciclu de fosforilare-defosforilare.
Na+, K+-ATP aza este cea mai studiata și are 3 roluri de baza:
Determina potențialul de membrana
Controleaza volumul celular
Intreține transportul cuplat cu gradiente ionice
Potențialul membranar determina ieșirea Cl- extracelular (ca si a Na+) este de 10 ori mai mare decat cea din citoplasma. Aceste diferențe in concentrații ionice echilibreaza concentrația mare a moleculelor organice intracelulare, egaland presiunile osmotice și prevenind intrarea apei.
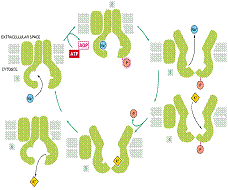
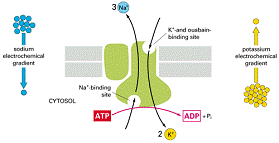
Model pentru pompa Na+, K+-ATP aza
Ca2+ -ATPaza
Pompa de calciu menține concentrația intracelulara a calciului la un nivel foarte scazut. In majoritatea celulelor animale și in eritrocite Ca2+ -ATPaza se afla in membrana plasmatica și are rolul de a transporta Ca2+ in afara celulei.
In celula musculara - Ca2+ -ATPaza e localizata in membrana RE și transporta ionii de calciu din citosol in interiorul RE (organit care are rolul de a depozita si concentra Ca2+; calsequestrina este o molecula proteica ce fixeaza 43 mol. Ca2+).
Eliberarea ionilor de calciu din RE catre citosol determina contracția, iar reintroducerea lor determina relaxarea musculara.
Ca2+ -ATPaza transporta cate 2 ioni de Ca2+ pentru fiecare molecula de ATP hidrolizata și necesita prezenta Mg2+.
H+-ATPaza lizozomala (ATP-azele din clasa V)
Sunt abundente in membrana lizozomala avand rolul de a menține interiorul acestor organite foarte acid (pH= 4,5-5) gradientul protonilor fiind mai mare fața de citosol de 100 x.
Se gasesc și in membranele unor celule acid-secretorii (exp. Celule epiteliale care marginesc vezica urinara la broasca).
ATP-azele membranare din clasa F
Se afla in membranele mitocondriale și este analoaga celor descrise mai sus, funcționeaza folosind un gradient de protoni, realizat de catre lanțul transportor de electroni din mitocondrie.
ATP sintetaza poate funcționa in ambele direcții: poate hidroliza ATP și prin aceasta pompeaza H+ prin membrana, sau poate sintetiza ATP, fiind pusa in funcțiune de fluxul de protoni ce are loc in sensulgradientului.
ATP sintetaza este responsabila pentru producția ATP din celula.
Unele sisteme de transport activ sunt intreținute de energia stocata in gradientul ionic, creat de catre pompele ionice ATP dependente și nu de energia eliberata de hidroliza ATP.
Exemplu: influxul de Na+ in citosol determinat de potențialul de membrana și de gradientul ionic, poate fi cuplat cu transportul moleculelor de glucoza, care va intra in celula impotriva gradientului de concentrație. Acest sistem, in care un ion, in deplasarea in sensul gradientului, determina transportul simultan al unei alte molecule (glucoza) impotriva gradientului, se numeste cotransport (simport = in acelasi sens, antiport = in direcții opuse).
Sistemul simport- asigura transportul glucozei si aminoacizilor din lumenul intestinal in celulele epiteliale ale acestuia și de aici in sange, transportul transmembranar este cuplat oligatoriu cu transportul a cate unui ion de Na+. Procesul se numeste transport transcelular. Gradientul de Na+ este o sursa de energie.
Sistemul antiport - este un proces prin care intrarea in celula a unui ion (exp: Na+) este cuplata obligatoriu cu ieșirea unei a doua molecule, contra gradientului de concentrație (exp: Ca2+).
Sistemul antiport Ca2+/ Na+
Funcționarea unui astfel de sistem antiport asigura reducerea contracțiilor miocardice. Na+, K+-ATP aza din membrana plasmatica a celulei cardiace creaza un gradient al concentrației de Na+ care susține exportul ionilor de Ca2+.
Medicamentele ouabaina și digoxina au o foarte mare semnificație clinica, ele crsc forța contracției miocardului in afecțiuni cardiace congestive. Efectul acestor medicamente consta in blocarea funcționarii Na+, K+-ATP aza ducand, in consecința, la creșterea concentrației intracelulare a ionilor de Na, scazand gradientul de sodiu, sistemul antiport Na+- Ca2+ funcționeaza mai puțin eficient, sunt exportați mai puțini ioni de calciu, iar concentrația intracelulara a Ca2+ crește, iar celulele cardiace se contracta mai frecvent și mai puternic.
Sistemul antiport de anioni (Cl-, HCO3 -)
In membrana eritrocitara proteina benzii 3 este un important antiport de anioni ce transporta in sens opus ioni negativi de clor și bicarbonat, transport esențial pentru funcționarea eritrocitului ca transportor de dioxid de carbon. Permeabilitatea benzii 3 la Cl- este de 100.000 de ori mai mare decat in alte celule.
CO2 dupa ce este eliberat de celule ajunge in capilarele sanguine, difuzeaza prin membrana eritrocitului, iar aici sub acțiunea anhidrazei carbonice, se transforma in anionul bicarbonic (H2O+CO2=H-+ HCO3-). Anionul bicarbonic este transportat la exteriorul eritrocitului de catre sistemul antiport , prin schimb cu un ion de Cl-.
Sistemul antiport Na+ - H+
Sistemul deține o importanța foarte mare in indepartarea excesului de protoni generați in metabolismul celulei animale. Asigura ridicarea ph in celule daca acesta scde sub limitele normale. Acest lucru este datorat unei proteine similare cu banda 3 din membrana eritrocitului, care expulzeaza bicarbonatul din citosol prin schimb cu Cl-.
Acest export de bicarbonat determina scaderea ph. Ph citosolic contribuie la activarea cailor metabolice necesare pentru creșterea și diviziunea celulara. (diferente mice exp: 7,4 la7,2 pot induce rapid stoparea mai multor funcții: sinteza ADN, sinteza ARN, catabolismul glucozei și sinteza proteinelor)
Sistemul antiport intalnit și la formarea acidului clorhidric de catre celulele parietale (celule oxintice) din stomac.
Imagine
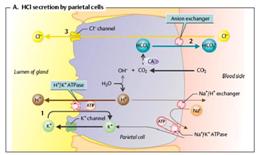
|
Politica de confidentialitate |
| Copyright ©
2024 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |