
STRUCTURA MOLECULARA A CROMOZOMILOR SI GENELOR
Unul dintre criteriile care stau la baza clasificarii organismelor este absenta sau prezenta nucleului individualizat. Celulele procariote lipsite de un veritabil nucleu, includ eubacteriile si archeobacteriile. De cealalta parte se afla celulele eucariote care poseda o membrana nucleara care separa cromozomii de restul celulei. Cromozomul unic al celulelor procariote este format dintr-o molecula de ADN dublu catenar circulara care contine cateva milioane de perechi de baze (Tabel 5.1). In general, genoamele eucariotelor au o marime si o complexitate variabila si contin mai multi cromozomi.
Atat celulele procariote cat si cele eucariote poseda mecanisme, pe de o parte comune si pe de alta parte distincte, pentru reglarea expresiei genelor si a fluxului de informatie de la ADN la proteine. Transcriptia ADN realizata de ARN polimeraze constituie prima etapa, comuna la toate organismele. La celulele procariote, ribozomii se fixeaza pe molecula de ARNm, in curs de sinteza, si realizeaza traducerea imediata a informatiei in proteine, in citoplasma. Din contra, la eucariote membrana celulara separa locul in care se realizeaza transcriptia de cel in care are loc traducerea. Transcriptul primar, ARNm imatur, contine o succesiune de secvente netraduse, numite introni, care vor fi eliminate prin procedeul de scindare a intronilor si sudare a exonilor ("splicing") in procesul de maturare a ARNm. In urma acestei etape, localizata in nucleu, ARNm trece in citoplasma unde este tradus. Maturarea ARNm prin "splicing" confera celulei eucariote posibilitatea diversificarii informatiei prin combinarea alternativa a exonilor, o alta etapa foarte importanta in dezvoltarea organismelor.
Organizarea genomului la procariote
Bacteriile, organismele procariote cele mai simple intalnite in toate mediile naturale, prezinta caracteristici comune. Adesea, un perete celular, format din peptidoglicani, inconjoara membrana plasmatica si confera celulei forma rigida. De asemenea, este prezenta o membrana externa care poseda un numar de flageli si pili variabil (Figura 5.1). Bacteriile au un compartiment citoplasmatic unic care contine un nucleoid format dintr-o molecula simpla circulara de ADN, ARN si proteine. Pe langa ADN prezent in nucleoid, citoplasma majoritatii bacteriilor contine mici molecule circulare de ADN, numite plasmide. Cel mai adesea acestea confera celulei rezistenta la antibiotice si la alte toxine. In conditii favorabile, bacteriile se multiplica rapid prin diviziune. Capacitatea lor de adaptare la conditiile de mediu si de diviziune rapida fac ca aceste celule sa constituie suporturi excelente pentru numeroase analize biochimice si de biologie moleculara, in vitro.
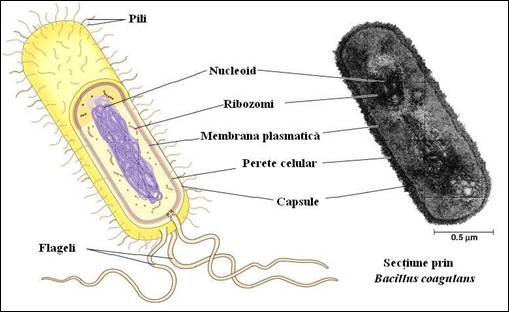
Figura 5. 1. Reprezentare clasica a unei celule procariote (dupa Campbell, et al., Biology, 2002)
Desi cea mai mare parte a genoamelor procariote sunt mai mici de 5 Mpb lungime, exista si cateva mai mari (Tabel 5.1). In prezentarea traditionala, genomul tipic procariotelor contine o singura molecula circulara de ADN, localizata la nivelul nucleoidului.
Aceasta abordare este adevarata pentru E. coli si pentru multe alte bacterii foarte studiate dar, specificam ca evolutia cunoasterii, in sfera genomului procariotelor, pune in discutie anumite concepte dezvoltate in era pre-genomica a microbiologiei. Aceste aspecte se refera atat la structura fizica a genomului procariotelor cat si la organizarea genetica.
Punctul de vedere traditional asupra cromozomului bacterian
Asemeni eucariotelor, genomul procariotelor trebuie sa se incadreze intr-un spatiu foarte restrans. Cromozomul circular de la E. coli are o circumferinta de 1,6 mm in timp ce dimensiunile celulei sunt de 1,0 x 2,0 μm. Structurarea necesara este realizata, ca si la eucariote, cu ajutorul proteinelor de legare la ADN ("DNA binding proteins") care compacteaza genomul intr-o forma organizata. Structura obtinuta nu prezinta similitudini substantiale cu cromozomul de la eucariote cu toate ca din punct de vedere didactic este folosita exprimarea de "cromozom bacterian".
Majoritatea cunostintelor pe care le avem despre organizarea ADN in nucleoid sunt rezultatul studiilor pe E. coli. Genomul circular al E. coli este suprarasucit. Suprarasucirile apar cand ture suplimentare sunt introduse sau indepartate din structura dublei elice (suprarasucire pozitiva/negativa. Suprarasucirea este modalitatea ideala de impachetare a unei molecule intr-un spatiu redus. Primele dovezi ca suprarasucirile sunt implicate in impachetarea genomului circular de la E. coli au fost obtinute in 1970 in urma examinarii unor nucleoizi izolati, fiind confirmate definitiv ca o trasatura a celulelor vii in 1981. Se considera ca, la E. coli supra-rasucirile sunt generate si controlate de doua enzime, ADN giraza si ADN topoizomeraza I.
Cea mai plauzibila explicatie privind compactarea nucleoidului este aceea ca ADN este atasat la proteine care restrang capacitatea de relaxare, astfel incat rotatia la nivelul unui situs de scindare are ca rezultat pierderea unei suprarasuciri dintr-un mic segment al moleculei. Modelul curent propune atasarea ADN de la E. coli pe structura unei proteine miez de la care radiaza 40-50 bucle in interiorul celulei. Fiecare bucla radiaza in interiorul celulei si contine aproximativ 100 kpb de ADN suprarasucit (Figura 5.2). Componentele proteice ale nucleoidului includ ADN giraza si ADN topoizomeraza I, cele doua enzime care sunt principale responsabile pentru mentinerea starii de suprarasucire, ca si un set de aproximativ patru proteine considerate a avea un rol mai specific in impachetarea ADN bacterian. Cea mai abundenta proteina este HU, care este foarte diferita structural de histonele eucariote dar actioneaza similar, formand un tetramer in jurul caruia se infasoara aproximativ 60 pb. In celula de E. coli sunt prezente aproximativ 60 000 copii ale HU care acopera aproximativ 1/50 din molecula de ADN, dar nu se cunoaste daca tetramerii sunt distribuiti de-a lungul ADN sau sunt pozitionati numai in regiunea centrala a nucleoidului.
Tabel 5. Marimea si complexitatea genomului la cateva virusuri si celule
|
Nume comun |
Marimea genomului (pb) |
Marimea relativa a genomului (E. Coli=1) |
|
|
Virusuri SV40 Bacteriofagul λ Virus herpes bovin |
5,2 x 103 4,8 x 104 1,4 x 105 |
1,3 x 10-3 1,2 x 10-2 3,5 x 10-2 |
|
|
Celule Procariote Mycoplasma capricolum Staphylococcus aureus Escherichia coli Myxococcus xanthus Bacillus megaterium |
Bacterii |
0,7 x 106 2,9 x 106 4,6 x 106 9,7 x 106 30 x 106 |
1,7 x 10-1 7,2 x 10-1 |
|
Celule Eucariote Ciuperci Saccharomyces cerevisiae Aspergillius niger Plante Arabidopsis thaliana Zea mays Alium cepa Animale Drosophila melanogaster Mus musculus Bos taurus Homo sapiens |
Drojdii Ciuperci Porumb Ceapa Musca Soarece Taur Om |
1,5 x 107 1,5 x 107 7,0 x 107 3,9 x 109 1,8 x 1010 1,8 x 108 2,7 x 109 3,2 x 109 3,4 x 109 |
1,7 x 101 9,7 x 102 4,5 x 103 4,5 x 101 6,7 x 102 8,0 x 102 8,5 x 102 |
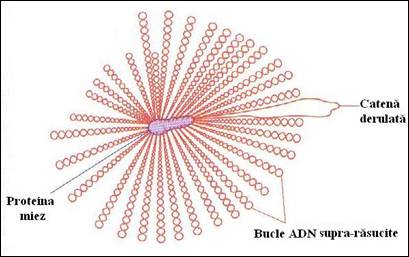
Figura 5. Model pentru structura nucleoidului de E. coli
Comentarii pe tema structurii nucleoidului procariot
In ultimii 10 ani a devenit foarte clar ca anatomia genomului procariot dezvoltata pe baza studiilor pe E. coli este foarte simplista. Desi marea majoritate a genoamelor bacteriene sau a cromozomilor de la archeobacterii sunt circulari, a fost descoperit un numar mare de forme lineare. Prima a fost descrisa pentru Borrelia burgdorferi, in 1989 de catre Ferdows si Barbour, urmatorii ani fiind realizate descoperiri similare pentru genul Streptomyces si alte bacterii.
Un alt aspect il reprezinta prezenta plasmidelor care reprezinta mici molecule de ADN, adesea dar nu intotdeauna circulare, care coexista cu cromozomul principal in celula bacteriana. Anumite tipuri de plasmide sunt capabile de integrare in genomul principal, in timp ce altele pot avea o existenta independenta. Plasmidele contin gene care, de obicei, nu se gasesc pe cromozomul principal, dar care sunt, in multe cazuri, neesentiale pentru bacterie, deoarece codifica pentru fenotipuri cum sunt rezistenta la antibiotice fara de care bacteriile pot trai in conditii de mediu normale (Tabel 5. 2). Multe plasmide sunt capabile de transfer de la o celula la alta, si aceleasi plasmide sunt adesea regasite in bacterii care apartin la diferite specii. Trasaturile diferite ale plasmidelor sugereaza ca ele reprezinta entitati independente care, in majoritatea cazurilor, nu sunt incluse in definitia genomului bacterian.
Pentru o bacterie de tipul E. coli, care are un cromozom de 4,6 Mpb si care poate contine variate combinatii de plasmide, nici una mai mare de cateva kilobaze, toate dispensabile, putem defini cromozomul principal ca "genom". In cazul altor procariote lucrurile nu stau la fel de simplu. De exemplu, cromozomul linear al Borrellia burgdoferi care are 910 kpb si contine 853 gene, este insotit de cel putin 17 plasmide circulare si lineare care impreuna contribuie cu alte 533 kpb si cu cel putin 430 gene. Desi, majoritatea acestor gene sunt orfane (fara functie identificata), au fost identificate si structuri indispensabile cum sunt genele pentru proteinele membranare si biosinteza purinelor. Se considera ca macar unele din cele 17 plasmide de la Borrelia sunt componente esentiale ale genomului. Astfel, este admisa posibilitatea ca anumite procariote sa posede genoame constituite din mai multe componente care includ un numar separat de molecule ADN. Aceasta situatie este mult mai apropiata de ceea care exista la eucariote decat la procariotele "tipice". De fapt, interpretarea informatiilor privind genomul de la Borrelia este inca controversata, si complicata datorita existentei unei bacterii inrudite, Treponema pallidum, al carei genom este format dintr-o singura molecula de ADN de 1138 kpb care contine 1041 gene, este lipsit de orice omologie cu genele prezente in plasmidele de la Borrelia.
Tabel 5. Trasaturile catorva plasmide tipice
|
Tip de plasmida |
Functiile genelor |
Exemple plasmide |
|
Rezistenta |
Rezistenta antibiotice |
Rbk la E. coli si alte bacterii |
|
Fertilitate |
Conjugare si transfer ADN intre bacterii |
F de la E. coli |
|
Ucigasa ("killer") |
Sinteza de toxine care ucid alte bacterii |
Col de la E. coli; produce colicina |
|
Degradativa |
Enzime pentru metabolismul moleculelor neobisnuite |
TOL pentru Pseudomonas putida; metabolism toluen |
|
Virulenta |
Patogenitate |
Ti de la Agrobacterium tumefaciens; capacitatea de a determina formarea galelor la dicotiledonate |
O alta complicatie privind structura fizica a genoamelor procariote este legata de diferentele dintre sistemele de impachetare pentru moleculele de ADN de la bacterii si archeobacterii. Una dintre motivatiile pentru care archeobacteriile sunt privite ca un grup distinct de organisme, diferite de bacterii, este faptul ca acestea nu poseda proteine de compactare de tipul HU, dar poseda proteine mult mai asemanatoare cu histonele. Cu toate ca, informatiile despre nucleoidul de la archeobacterii sunt destul de incomplete, se presupune ca proteinele asemanatoare histonelor au un rol important in impachetare.
Cunoastem deja ca genomul bacterian are o structura genetica compacta cu foarte putin spatiu intre gene. Aceasta organizare este foarte bine demonstrata si tipica la genomul de la E. coli. La acest organism ADN care nu codifica pentru diferite gene reprezinta numai 11% din total si este distribuit pe toata lungimea sub forma unor segmente mici. O serie de teorii sustin ca organizarea compacta este benefica pentru replicarea rapida a genomului cu toate ca ipotezele nu pot fi sustinute de dovezi experimentale.
Operonii sunt caracteristici ale genoamelor procariote
Cea de a doua caracteristica a genomului de la procariote, ilustrata la E. coli, este prezenta operonilor. Operonii reprezinta un grup de gene localizate una langa cealalta. Toate genele din structura unui operon sunt exprimate ca o singura unitate. Acest tip de aranjament este comun genoamelor procariote si poate fi ilustrat la E. coli cu operonul lactozei care a fost descoperit in 1961 de catre Jacob si Monod. Acesta contine trei gene implicate in transformarea dizaharidului lactoza in unitatile sale componente (glucoza si galactoza). Genele operonului lactoza sunt implicate in utilizarea lactozei ca sursa de energie de catre E. coli. Deoarece, lactoza nu este o componenta comuna a mediul inconjurator pentru E. coli, majoritatea timpului operonul nu este exprimat si enzimele pentru utilizarea lactozei nu sunt sintetizate de bacterie. Cand lactoza devine disponibila se realizeaza activarea operonului si cele trei gene sunt exprimate impreuna. Operonul lactoza reprezinta un exemplu clasic de reglare a expresiei genelor la bacterii (Figura 5.3 a
La E. coli si la Bacillus subtilis exista aproximativ 600 operoni. In majoritatea cazurilor, genele unui operon sunt corelate functional si codifica pentru un set de proteine implicate intr-o anumita cale metabolica, cum ar fi utilizarea unui glucid ca sursa de energie sau sinteza unui aminoacid (operon triptofan, Figura 5.3 b). La alte bacterii sau archeobacterii exista posibilitatea ca operonii sa contina gene implicate in diferite cai metabolice. Este cazul unuia dintre operonii de la Aquiflex aeolicus care contine sase "gene linkate" care codifica pentru: i) doua proteine implicate in recombinare; ii) o enzima folosita in sinteza proteica; iii) o proteina necesara pentru mobilitate; iv) o enzima utilizata in sinteza nucleotidica; v) o enzima pentru sinteza lipidica (Figura 5.3c). In concluzie, teoria ca expresia unui operon conduce la sinteza coordonata a mai multor enzime necesare pentru o singura cale metabolica nu este valabila pentru toate speciile.
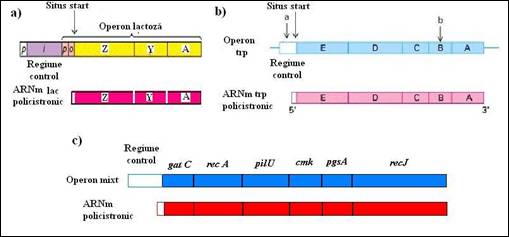
Figura 5. Diferite tipuri de operoni: a) operonul lactoza de la E. coli; b) operonul triptofan de la E. coli; c) operonul mixt de la Aquifex aeolicus
Continutul minim de gene si identitatea genelor specifice
Cu toate ca numarul de genoame procariote secventializate integral creste continuu, nu este inca posibila realizarea unui catalog complet cu continutul genelor fiecarei specii, din simplul motiv ca functiile anumitor gene raman necunoscute. De exemplu, genomul de la E. coli contine 4 288 gene care codifica pentru proteine dar peste 1 500 din acestea sunt orfane. Chiar daca informatia este adesea incompleta, este interesant sa examinam rolul genelor ale caror functii sunt cunoscute, si sa apreciem numarul diferit al genelor implicate in diferite activitati biochimice la o bacterie de tipul E. coli (Tabel 5.3
Cataloagele de gene sunt mult mai interesante cand comparatiile se realizeaza intre diferite specii. De exemplu, la E. coli 243 de gene sunt implicate in metabolismul energetic, la Haemophylus influenzae numai 112 si la Mycoplasma genitalium numai 31. Aceste comparatii pot duce la speculatii privind numarul minim de gene necesar pentru supravietuirea unei celule. Majoritatea demersurilor realizate in acest sens au demarat cu studiul celor mai mici genoame, cel al Mycoplasma genitalium si cel a Mycoplasma pneumoniae, care contin numai 470 si respectiv 679 gene. Consideratii teoretice au dus la sugestia ca 256 gene reprezinta minimul necesar, dar experimentele de inducere de mutatii sugereaza ca pentru Mycoplasma sunt necesare minim 300. Studii similare interesante au fost realizate pentru genele specifice care determina distinctia dintre o specie si alta.
Tabel 5. Catalog comparativ de gene pentru E. coli, H. influentze, M. genitalium
|
Categorie |
Numar de gene |
||
|
E. coli |
H. influentzae |
M. genitalium |
|
|
Total gene pentru proteine | |||
|
Biosinteza aminoacizi | |||
|
Biosinteza cofactori | |||
|
Biosinteza nucleotide | |||
|
Proteina anvelopa celulara | |||
|
Metabolism energetic | |||
|
Metabolism intermediar | |||
|
Metabolism lipidic | |||
|
Replicare ADN, recombinare, reparare | |||
|
Pliere proteine | |||
|
Proteine reglatoare | |||
|
Transcriptie | |||
|
Translatie | |||
|
Preluarea de molecule din mediu | |||
Din cele 470 gene ale genomului de la M. genitalium, 350 sunt prezente si la bacterii mai indepartate evolutiv, cum este Bacillus subtilis. Aceasta sugereaza ca trasaturile biochimice si structurale care disting Mycoplasma de Bacillus sunt codificate in aproximativ cele 120 gene care sunt unice la fiecare tip. Din nefericire, astfel de comparatii privind genele particulare nu dau raspunsuri cheie privind caracteristicile care individualizeaza fiecare tip de microorganism.
Structura materialului genetic la eucariote
In ciuda diferentelor evidente de forma si de functie, diferitele tipuri celulare care compun un organism multicelular, fie ca este vorba de o ciuperca sau de un mamifer, contin un lot complet de gene. Vazuta din exterior o celula din constitutia unui cartilaj si un neuron nu se aseamana deloc. Cu toate acestea cele doua tipuri celulare contin acelasi complement de gene.
Informatia genetica, prezenta intr-o celula eucariota specializata, se poate compara cu o carte care contine toate informatiile necesare constructiei unei cladiri cu utilitati multiple. In cursul realizarii constructiei, sunt probabil necesare toate planurile dar numai o mica parte a informatiei va trebui consultata in timpul activitatii de constructie a unui etaj sau a unei singure camere. Aceasta abordare este adevarata si pentru un ovul fecundat, care contine ansamblul instructiunilor genetice, si care este fidel replicat si informatia distribuita tuturor celulelor organismului in dezvoltare. In consecinta, celulele poarta mult mai multa informatie genetica decat pot utiliza vreodata. Acestea poseda mecanisme care le permit sa exprime informatia genetica, in mod selectiv, urmand numai instructiunile care privesc numai o singura celula intr-un moment particular al existentei acesteia. In acest subcapitol, o sa analizam modalitatile care le permit celulelor eucariote sa structureze totalitatea materialului genetic pe care il contin, in conditiile controlului coordonat al expresiei genelor menite sa asigure numai sinteza anumitor proteine. Mai intai vom descrie structura si proprietatile nucleului celulei eucariote, care contine majoritatea informatiei genetice si mecanismele de reglare a acesteia.
Nucleul celulei eucariote
In ciuda importantei sale pentru stocarea si utilizarea informatiei genetice, nucleul celulei eucariote poseda o morfologie destul de comuna (Figura 5.4). Continutul nuclear reprezinta o masa vascoasa amorfa de materie inchisa intr-un invelis nuclear complex. In nucleul unei celule tipice, in interfaza, se evidentiaza: i) cromozomii, prezenti sub forma unor fibre nucleoproteice foarte alungite, cromatina; ii) matricea nucleara care este formata dintr-o retea fibrilara care contine proteine; iii) unul sau mai multi nucleoli, structuri amorfe, opace la microscopie electronica care sunt sediul sintezei ARN ribozomal si al asamblarii ribozomilor; iv) nucleoplasma, substanta lichida care contine toate componentele si elementele nucleului eucariot. In figura 5.4 sunt ilustrate schematic diferitele elemente ale nucleului eucariot.
Invelisul nuclear
In opozitie cu termenul de membrana nucleara, invelisul nuclear desemneaza structura complexa care se afla la frontiera dintre nucleul si citoplasma unei celule eucariote. Intr-o celula, separarea materialului genetic si a citoplasmei care il inconjoara este poate caracteristica cea mai importanta care distinge eucariotele de procariote. Aparitia invelisului nuclear este cu siguranta un punct de reper in evolutia biologica. Invelisul nuclear consta in mai multe elemente distincte (Figura 5.4). Continuitatea invelisului care delimiteaza nucleul este asigurata de doua membrane concentrice separate de un spatiu intermembranar de 10 la 50 nm. Cele doua membrane sunt fuzionate din loc in loc si formeaza pori circulari care sunt compusi dintr-un ansamblu complex de proteine. Densitatea porilor nucleari variaza de la aproximativ 2 la 4 pe mm2 in invelisul nuclear al eritrocitelor de pasare, relativ inactiv din punct de vedere metabolic, la mai mult de 60 mm2 in cel al ovocitelor care se pregatesc sa raspunda nevoilor viitoare ale dezvoltarii embrionare.
O celula de mamifere, de marime medie, poseda aproximativ 3 000 de pori nucleari. Membrana externa este in general presarata cu ribozomi si adesea in continuitate cu membrana reticulului endoplasmatic (Figura 5.4
Suprafata interna a invelisului nuclear este bordata de o plasa fibrilara densa, numita lamina nucleara. In anumite celule, cum este ovocitul de amfibian, aceasta lamina formeaza un strat destul de continuu, in timp ce in altele pare mult mai fragmentata. Consideram ca lamina nucleara reprezinta un suport structural pentru invelisul nuclear si serveste la fixarea fibrelor de cromatina la periferia nucleului (Figura 5.4
Fibrilele laminei nucleare au un diametru de aproximativ 10 nm si sunt compuse din polipeptide numite lamine, care apartin aceleiasi superfamilii de polipeptide care compun filamentele intermediare ale citoscheletului. Ca si pentru componentele intermediare ale citoplasmei, integritatea laminei nucleare este controlata de catre procesul de fosforilare/defosforilare. Dezasamblarea laminei nucleare, inainte de mitoza, este indusa de catre fosforilarea laminelor de catre o kinaza specifica.
Structura si functia complexului porilor nucleari
Invelisul nuclear este o bariera care separa doua din cele mai importante compartimente ale celulei, nucleul si citoplasma, porii fiind porti in aceasta bariera. Contrar membranei plasmatice, care impiedica trecerea macromoleculelor intre citoplasma si spatiul extracelular, anvelopa nucleara este un centru activ pentru ARN si proteinele care se deplaseaza in cele doua sensuri, intre compartimentele pe care aceasta le separa. Replicarea si transcriptia materialului genetic, in nucleu, necesita participarea unui mare numar de proteine care se sintetizeaza in citoplasma si sunt transportate prin invelisul nuclear. Invers, moleculele de ARNm, ARNt si subunitatile ribozomilor care sunt fabricate in nucleu trebuie sa fie transportate prin membrana nucleara, in sens invers.
Anumite elemente, cum sunt moleculele ARNsn (ARN "small nuclear"), se deplaseaza in cele doua directii. Acestea sunt sintetizate in nucleu, se asambleaza in particule ribonucleoproteice (RNP), in citoplasma, si sunt reimportate in nucleu unde intervin in maturarea ARNm. Acest trafic intens se realizeaza prin porii nucleari. Tinand seama ca particule materiale, de talia subunitatilor ribozomale, pot sa treaca prin porii nucleari, putem presupune ca acestia sunt canale deschise dar, de fapt, este exact contrar. Porii nucleari contin un aparat complex, in forma de cos, numit Complexul Porului Nuclear (CPN) care obtureaza porul ca un dop si care se reliefeaza in afara in citoplasma si in nucleoplasma. Structura CPN este prezentata in modelul schematic din figura 5.5 si in micrografiile electronice din figura 5.6 .
CPN este un enorm complex molecular cu simetrie octogonala care se estimeaza ca are in structura 100 la 200 polipeptide. Masa CPN este cam de 30 ori mai mare decat masa unui ribozom. Structura moleculara a complexului CPN a fost determinata de-a lungul timpului gratie tehnicilor de microscopie electronica si de analiza de imagini din ce in ce mai performante. Daca sunt injectate solutii, ale moleculelor cu masa moleculara mica, in citoplasma unei celule acestea pot penetra rapid in porii nucleari prin simpla difuzie. Experimentul sugereaza ca aceste substante sunt capabile sa treaca printre fantele existente intre razele cosului.
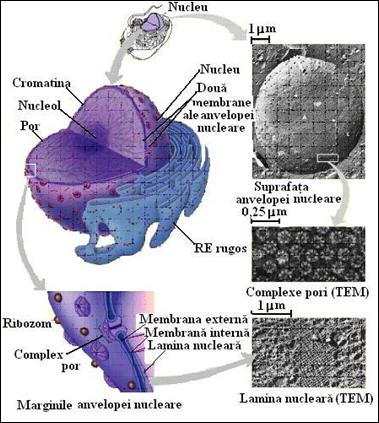
Figura 5.4 Nucleul celular si caracteristicile sale (dupa Campbell, et al., Biology, 2002)
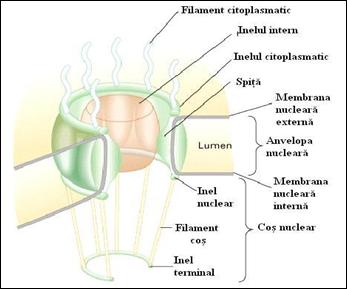
Figura 5. Complexul porului nuclear, model schematic
(dupa Molecular Cell Biology Lodish H., et. al., 2000)
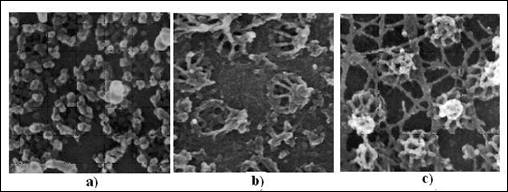
Figura 5.6. Anvelopa nucleara a ovocitelor de Xenopus vizualizata prin microscopie electronica: a) fata citoplasmatica a complexului porului nuclear (CPN); b) fata nucleoplasmatica a anvelopei nucleare dupa indepartarea membranei nucleare a CPN, evidentiind structura sub forma de cos; c) fata nucleoplasmatica a anvelopei nucleare dupa indepartarea membranei nucleare prin tratament bland cu detergent. Reteaua laminara, care se insereaza in inelul nuclear al NPC, este expusa dupa acest tratament (dupa Molecular Cell Biology Lodish H., et. al., 2000)
Capacitatea moleculelor mai mari (proteine si nucleoproteine) de a trece din citoplasma in nucleu depinde de sediul lor obisnuit de rezidenta, in nucleu sau in afara acestuia. De exemplu, daca o proteina citoplasmatica, cum este albumina serica bovina, este marcata radioactiv si injectata in citoplasma, ea are tendinta de a ramane acolo. Din contra, daca injectarea este realizata cu o proteina cu localizare nucleara, nucleoplasmina, proteina marcata va fi regasita imediat in nucleu.
In 1982, Robert Laskey si colaboratorii sai de la Medical Research Council, Anglia, au descoperit ca nucleoplasmina, una dintre cele mai abundente proteine nucleare din ovocitele de amfibieni, contine o secventa de aminoacizi in apropierea extremitatii C terminale care functioneaza ca un Semnal de Localizare Nucleara (SLN), care ii permite sa treaca prin porii nucleari si sa intre in nucleu. De atunci au fost identificate o serie de alte SLN, cea mai mare parte a acestora fiind constituite din scurte secvente de aminoacizi cu sarcina pozitiva. In principiu, orientarea (adresarea) proteinelor catre nucleu este asemanatoare cu transportul altor proteine care sunt distribuite in anumite organite, de tipul mitocondriei sau peroxizomilor. In toate cazurile, proteinele poseda o "eticheta" specifica care este recunoscuta de catre un receptor particular de la suprafata organitului tinta.
Importul proteinelor in nucleu trece prin mai multe etape: i) proteina care trebuie importata se cupleaza cu un receptor pentru SLN care pare a fi localizat la suprafata filamentelor departate de inelul extern al porului catre citoplasma (Figura 5. 7); ii) interactia intre proteina nucleara si receptorul pentru SLN permite acostarea proteinei pe fata citoplasmatica a complexului porului nuclear prin formarea unui complex de transport; iii) deplasarea complexului de transport prin por necesita energie eliberata din hidroliza ATP. Aceasta implica o modificare a conformatiei transportorului, structura mare in forma de dop (inel intern) situata in centrul complexului porului nuclear (Figura 5.5). Modificarea de conformatie deschide un canal in centrul transportorului si permite moleculelor situate in proximitatea acestuia sa patrunda in nucleoplasma. Dupa acelasi principiu, substantele care trec din nucleu in citoplasma declanseaza, probabil, deschiderea transportorului in sens invers.
In figura 5.7 este prezentat un model schematic de import nuclear al unei proteine (cargo) dupa cuplarea acesteia cu semnalul de localizare nucleara (SLN). In citoplasma, importinele si interactioneaza cooperativ cu proteina cargo care urmeaza a fi transportata, prin legarea importinei la SLN. Subunitatea importinei , din complexul cargo trimeric rezultat, interactioneaza cu componentele CPN, translocand complexul in nucleoplasma prin mecanisme destul de putin cunoscute. Aceasta translocare necesita hidroliza ATP. In nucleoplasma, Ran-GTP interactioneaza cu importina , determinand disocierea complexului cargo si eliberand proteina transportata in nucleoplasma. Pentru a sustine un alt ciclu de import, importina monomera si complexul importina -Ran-GTP sunt transportate inapoi in citoplasma. Proteina de activare RanGAP (Ran GTP activating protein), din citoplasma, stimuleaza transformarea Ran-GTP la Ran-GDP avand drept rezultat o transformare conformationala la nivelul Ran care determina disocierea de importina . Importina libera interactioneaza apoi cu importina si se formeaza un nou complex cargo purtator al unui semnal bazic SLN, care initiaza un alt ciclu de import nuclear. Se presupune ca Ran-GDP este si el translocat prin porii nucleari din citoplasma in nucleoplasma, unde factorul nucleotidic de schimb, Ran (RCC1), determina eliberarea GDP si relegarea GTP.
Un por nuclear individual este capabil sa importe proteine si sa exporte ARN si ribonucleoproteine. Acest transport bidirectional poate fi vizualizat prin microscopie electronica. De asemenea, tendinta de acumulare in unul dintre cele doua compartimente celulare a unor proteine purtatoare de semnale de localizare in citoplasma sau in nucleu poate fi vizualizata prin microscopie de fluorescenta (Figura 5.8) sau confocala.
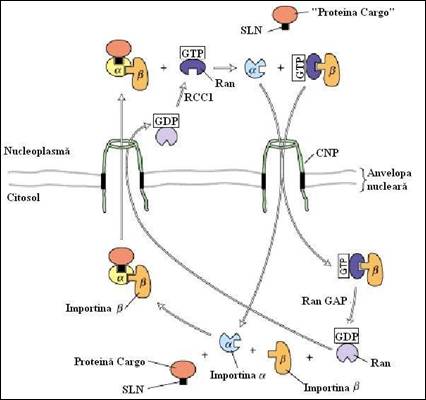
Figura 5.7. Mecanismul propus pentru transportul, din citoplasma in nucleu, a proteinelor "cargo"care contin un semnal de localizare nucleara (SLN) (dupa Molecular Cell Biology Lodish H., et. al., 2000).
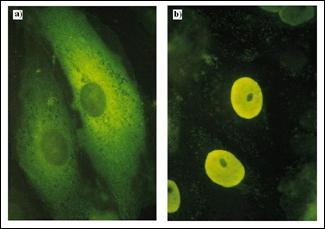 Figura 5.8. Semnalul
de Localizare Nucleara (SLN) al
antigenului T (SV40) poate directiona o proteina citoplas-matica
in nucleu: a) Piruvat kinaza normala, localizata in citoplasma,
vizualizata prin imunofluorescenta dupa tratarea celulelor
in cultura cu un anticorp specific; b) piruvat kinaza himera, care
contine un semnal SNL al SV40 la capatul sau N-terminal, este
directionata catre nucleu. Proteina himera a fost
exprimata in urma transfectiei unei gene modificate produsa prin
fuziunea unui fragment al unei gene virale care codifica NLS al SV40 cu
gena piruvat kinazei (dupa Molecular Cell Biology Lodish H., et.
al., 2000).
Figura 5.8. Semnalul
de Localizare Nucleara (SLN) al
antigenului T (SV40) poate directiona o proteina citoplas-matica
in nucleu: a) Piruvat kinaza normala, localizata in citoplasma,
vizualizata prin imunofluorescenta dupa tratarea celulelor
in cultura cu un anticorp specific; b) piruvat kinaza himera, care
contine un semnal SNL al SV40 la capatul sau N-terminal, este
directionata catre nucleu. Proteina himera a fost
exprimata in urma transfectiei unei gene modificate produsa prin
fuziunea unui fragment al unei gene virale care codifica NLS al SV40 cu
gena piruvat kinazei (dupa Molecular Cell Biology Lodish H., et.
al., 2000).
Nucleul este un organit organizat
Specialistii in biologie celulara se bazeaza mult pe studiile biochimice pentru a intelege activitatile nucleului. Cu toate ca acestea furnizeaza un numar mare de informatii, distrug toate structurile moleculare organizate care pot exista in nucleu si dau impresia ca acesta nu este decat un "sac" de elemente repartizate la intamplare. In acelasi timp, o serie de studii de microscopie ne fac sa realizam ca nucleul este in realitate un compartiment foarte organizat. De exemplu, fibrele de cromatina, care compun un cromozom interfazic, nu sunt dispersate in tot nucleul, cum am putea crede, ele sunt concentrate intr-un domeniu specific care nu se suprapune deloc cu domeniile altor cromozomi.
Interactiunile intre extremitatile cromozomilor si anvelopa nucleara reprezinta un alt mod de organizare a cromozomilor in nucleu. Aceasta asociere este evidenta mai ales in meioza, cand cromozomii pot sa se indeparteze de anvelopa nucleara si sa formeze un "buchet".
De mai bine de treizeci de ani se stie ca ARN ribozomal este sintetizat in nucleol, dar se considera ca celelalte molecule de ARN, si mai ales ARNm se asamblau in tot nucleul. Cercetarile ultimilor zece ani au demonstrat clar ca moleculele de ARNm sunt sintetizate intr-un numar limitat de situsuri discrete situate in intregul nucleu. Daca sunt identificate cu ajutorul unor sonde marcate fluorescent, situsurile de sinteza si de maturare a moleculelor de pre-ARNm apar sub forma unor "pete" stralucitoare in nucleoplasma nemarcata (Figura 5.9 b,c). Fiecare din cele 20 la 50 de pete observate intr-un anumit nucleu reprezinta situsul sintezei mai multor molecule de ARNm diferite. Diferitele elemente ale nucleului sunt organizate in acest compartiment printr-o retea interactiva complexa de filamente care compun matricea nucleara.
Matricea nucleara
Cand nucleele izolate sunt tratate cu detergenti anionici si o concentratie crescuta de saruri (NaCl 2M), pentru a extrage lipidele si aproape toate proteinele histonice si nehistonice ale cromatinei, ADN apare ca un halou care inconjoara un centru nuclear rezidual (Figura 5.10a). Daca fibrele de ADN sunt apoi digerate cu nucleaza, ramane o structura care pastreaza forma nucleului de origine, dar ea este compusa din fibrile proteice subtiri care se incruciseaza pe suprafata spatiului nuclear (Figura 5.10b). Aceasta retea fibrilara insolubila este matricea nucleara.
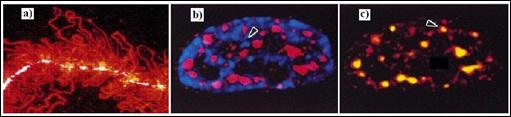 Figura 5.9. Localizarea situsurilor de sinteza si de maturare a pre-ARN cu
sonde fluorescente: a) vizualizarea procesului de transcriptie prin
colorare cu DAPI pentru evidentierea genei si cu anticorpi
fata de heterogen nuclear ribonucleo proteine, marcati cu
fluoresceina; b) evidentierea procesului de poliadenilare prin
colorare cu poli-dT marcata cu rodamina si cu DAPI pentru
evidentierea ADN; c) evidentierea "splicingului" prin utilizarea de
anticorpi monoclonali, fata de proteina de splicing SC-35, marcati
cu fluoresceina (dupa Molecular Cell Biology Lodish H., et.
al., 2000)
Figura 5.9. Localizarea situsurilor de sinteza si de maturare a pre-ARN cu
sonde fluorescente: a) vizualizarea procesului de transcriptie prin
colorare cu DAPI pentru evidentierea genei si cu anticorpi
fata de heterogen nuclear ribonucleo proteine, marcati cu
fluoresceina; b) evidentierea procesului de poliadenilare prin
colorare cu poli-dT marcata cu rodamina si cu DAPI pentru
evidentierea ADN; c) evidentierea "splicingului" prin utilizarea de
anticorpi monoclonali, fata de proteina de splicing SC-35, marcati
cu fluoresceina (dupa Molecular Cell Biology Lodish H., et.
al., 2000)
Matricea nucleara nu este numai un schelet care mentine forma nucleului sau un esafodaj pe care se organizeaza buclele de cromatina; ea serveste si de fixare pentru mecanismele care intervin in diferite activitati ale nucleului, cum sunt transcriptia, maturarea ARN si replicarea. De exemplu, daca celulele sunt incubate in prezenta de precursori radioactivi ai ARN sau ADN in timpul unei scurte perioade, gasim ca aproape toti acizii nucleici sintetizati sunt asociati fibrilelor matricei nucleare.
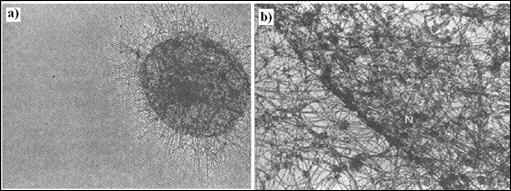 Figura 5.10. Matricea
nucleara: a) micrografie electronica a unui nucleu izolat in
prezenta de detergenti si saruri 2M; b) micrografie
electronica a unei regiuni de fibroblast de soarece in urma tratamentului
cu detergent si inlaturare a cromatinei si ADN
Figura 5.10. Matricea
nucleara: a) micrografie electronica a unui nucleu izolat in
prezenta de detergenti si saruri 2M; b) micrografie
electronica a unei regiuni de fibroblast de soarece in urma tratamentului
cu detergent si inlaturare a cromatinei si ADN
Proba cea mai clara privind importanta matricei nucleare a fost furnizata de o serie de cercetari efectuate in laboratorul lui Jeanne Lawrence de la Universitatea din Massachusetts. Utilizand hibridizarea fluorescenta, in situ, pentru a identifica localizarea secventelor specifice de ADN sau ARN. Lawrence si colaboratorii sai au constatat ca moleculele de ARN transcrise, pornind de la o anumita gena, apar ca o trena intr-o serie limitata de nuclee. Trena fluorescenta apare datorita prezentei mai multor sute de molecule de pre-ARNm transcrise pornind de la o singura gena (Figura 5.9a). Concentrarea transcriptilor intr-un loc limitat ne sugereaza ca acestia nu sunt liberi sa difuzeze plecand de la ADN de pe care au fost transcrisi. Se pare ca acestia sunt mentinuti locului de unele dintre elementele nucleului pe toata perioada maturarii lor.
Daca nucleele sunt tratate cu detergenti si cu saruri concentrate, practic toate moleculele de pre-ARNm care au fost transcrise raman asociate matricei nucleare reziduale. De fapt, urmele de ARNm fluorescente care au fost observate inainte de extractia dintr-un anumit nucleu isi pastreaza exact pozitia si dupa extractie, deoarece moleculele de ARNm nou formate sunt asociate elementelor matricei celulare. De asemenea, orientarea trenelor sugereaza ca moleculele de ARN parasesc locurile de sinteza pentru a se localiza in situsuri mai apropiate de periferia nucleului. In cursul deplasarii moleculelor de ARNm fata de gena de la care sunt sintetizate, intronii sunt eliminati din structura transcriptilor prin mecanismul de "splicing". Orientarea trenei, dinspre interiorul nucleului catre periferia sa, este compatibila cu un model in care matrita nucleara ghideaza moleculele de ARN, in curs de maturare, de la locul in care sunt transcrise catre porii nucleari pentru a iesi din nucleu.
Cromozomii
Cromozomii se individualizeaza structural la inceputul mitozei si "par sa dispara" din nou la sfarsitul acesteia. Acestia sunt constituiti dintr-un complex de ADN si proteine numit cromatina. Cromatina exista in diverse stari in diferite faze ale ciclului celular. Cromozomii sunt mult mai scurti decat lungimea moleculelor de ADN pe care le contin. In medie, un cromozom uman contine o molecula de ADN de aproximativ 5 cm. Compactarea acestui material genetic, in structura cromozomului, presupune existenta unui sistem organizat de impachetare care sa permita totusi functionarea genomului si exprimarea genelor. Observate la microscopul electronic, cea mai mare parte a organitelor intracelulare prezinta o structura care ne ofera informatii interesante despre functia lor. Totusi, micrografiile electronice ale preparatelor fine ale nucleului ofera destul de putine informatii privind natura cromozomilor in interfaza. In aceasta etapa, cromozomii sunt mult mai putin vizibili deoarece cromatina se afla intr-o stare mai relaxata (Figura 5.11a
Structura cromozomului mitotic
Dispersarea cromatinei in celulele in interfaza face mult mai usoara replicarea si transcriptia ADN. Din contra, cromatina celulelor mitotice este sub forma cea mai condensata, care usureaza "livrarea" unui pachet intact de ADN fiecarei celule fiice. Cromozomii mitotici sunt foarte utili, atat pentru biologi cat si pentru medici, deoarece ei contin un lot complet de material genetic celular si pot fi pusi in evidenta prin tehnici simple (Figura 5.11b). Cand un cromozom se condenseaza in timpul profazei mitotice, el adopta o forma distincta si constanta determinata, in principal, de lungimea moleculei de ADN si de pozitia centromerului. Se pot evidentia cromozomii mitotici ai unei celule in diviziune prin tehnica descrisa in figura 5. 12a. In aceasta tehnica, celulele sunt sparte dupa oprirea diviziunii celulare in mitoza in urma folosirii colchicinei. Cromozomii mitotici sunt fixati la suprafata unei lame unde ocupa o suprafata foarte mica si colorati cu diferite tipuri de reactivi (Tabel 5. 3). De exemplu, tehnica in care se realizeaza o proteoliza limitata si colorarea cu reactiv Giemsa poarta numele de "bandare G". Prin aceasta se evidentiaza prin benzi inchise la culoare regiunile bogate in AT si prin benzi pale cele bogate in GC.
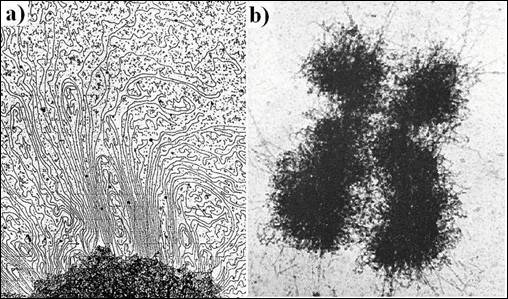
Figura 5.11. Comparatie intre starea cromatinei in interfaza si a cromatinei compactate in mitoza: a) micrografie electronica prezentand starea dispersata a cromatinei in interfaza; b) micrografie electronica a unui cromozom mitotic (dupa Molecular Cell Biology Lodish H., et. al., 2000)
Dupa fotografiere cromozomii individuali sunt decupati si aranjati in perechi. Se realizeaza un cariotip in care cromozomii omologi sunt dispusi in ordinea descrescatoare a marimii asa cum se prezinta in figura 5.12b. Celulele somatice de la om contin 46 cromozomi care pot fi grupati in 22 de perechi omologe si cromozomii sexuali care sunt XX la femei si XY la barbati. Studiul cariotipului este o tehnica de baza pentru citogeneticieni. Cu colorantul Giemsa se obtin intre 400 si 800 benzi pentru un set haploid de cromozomi. Preparatele de cromozomi mitotici sunt curent realizate pornind de la culturi de celule sanguine pentru a identifica indivizii purtatori de anomalii cromozomale. Exista posibilitatea determinarii cromozomilor suplimentari, absenta lor sau o serie de alte modificari vizibile la microscopul optic.
O alta modalitate de prelucrare o reprezinta colorarea cu quinacrina (bandarea Q) care evidentiaza striuri transversale. Deoarece distributia benzilor Q este caracteristica, fiecarui cromozom de la fiecare specie, tehnica permite identificarea rapida a cromozomilor si realizarea unor comparatii intre specii. De asemenea, se pot detecta anomalii minime din structura cromozomului prin identificarea modificarilor care apar in repartitia benzilor. Benzile Q stralucitoare contin, de obicei, secvente de ADN bogate in A-T si un numar redus de gene care codifica pentru proteine.
Bratul scurt al cromozomului este desemnat cu litera p ("petit"), iar bratul lung cu litera q (urmatoarea litera din alfabet). In tehnicile de bandare, fiecare brat al cromozomului este impartit in doua sau mai multe regiuni prin benzi mari. Regiunile sunt numerotate cu 1, 2, 3 pornind de la centromeri catre capete. Fiecare regiune este divizata in benzi si subdiviziuni ale benzilor. De exemplu, Xp21.2 se refera la segmentul cromozomial localizat pe bratul scurt al cromozomului X, in regiunea 2, banda 1 si sub-banda 2.
O alta tehnica, mult mai recenta, dar care a intrat in setul analizelor de de rutina pentru stabilirea cariotipului, este hibridizarea fluorescenta, in situ - FISH ("Fluorescence In Situ Hybridization"). Metoda prezinta o limitare tehnica pentru stabilirea cariotipului deoarece nu poate fi aplicata decat celulelor in diviziune sau celor la care diviziunea este indusa, in vitro. Aceasta problema poate fi depasita daca se folosesc sonde ADN care recunosc secvente specifice de pe cromozomi. Astfel de sonde, marcate fluorescent, sunt folosite pentru tratarea nucleelor interfazici. Sondele se leaga la secventele complementare de pe cromozomi si marcheaza specific cromozomii care pot fi vizualizati prin microscopie de fluorescenta. Astfel, FISH poate fi folosita pentru identificarea cromozomilor din nucleii in interfaza. Aplicatiile nu se limiteaza numai la acest aspect. Prin folosirea unor sonde specifice, pentru regiuni bine definite de pe cromozom, FISH se foloseste pentru identificarea microdeletiilor fine si translocarilor complexe care nu pot fi detectate prin tehnici clasice. De asemenea, FISH reprezinta o metoda de cartare a genelor, de interes clinic, nou izolate. O varianta extinsa a tehnicii FISH, numita "Chromosome Painting", permite identificarea tuturor cromozomilor unei celule prin folosirea unor seturi de sonde marcate fluorescent care recunosc secvente particulare de-a lungul cromozomilor. Numarul cromozomilor care pot fi detectati simultan poate fi limitat numai de disponibilitatea colorantilor fluorescenti care pot fi excitati la diferite lungimi de unda. De exemplu, tehnica nu poate fi folosita pentru vizualizarea simultana a tuturor celor 46 de cromozomi de la om. Aceasta limita a fost depasita prin introducerea "cariotipului spectral" care presupune folosirea unor combinatii de cinci fluorocromi si semnale adecvate controlate de computer. In acest fel este posibila vizualizarea fluorescenta a tuturor cromozomilor (Figura 5.13 b
Tabel 5.4. Tehnici de colorare folosite pentru bandarea cromozomilor
|
Tehnica |
Procedeu |
Tipul benzilor |
|
Bandarea G |
Proteoliza limitata, colorare Giemsa |
Benzi intunecate bogate in AT; Benzi pale bogate in GC |
|
Bandarea R |
Denaturare prin caldura, colorare Giemsa |
Benzi intunecate bogate in GC Benzi pale bogate in AT |
|
Bandare Q |
Colorare cu quinacrina |
Benzi intunecate bogate in AT Benzi pale bogate in GC |
|
Bandare C |
Denaturare cu hidroxid de bariu, colorare Giemsa |
Evidentierea de benzi intunecate de heterocromatina |
Centromeri
Se poate remarca ca toti cromozomii reprezentati in figurile 5.11 la 5.13 poseda o regiune unde suprafata lor exterioara este net rascroita. Aceasta scobitura care este vizibila foarte bine in figura 5.11b reprezinta centromerul cromozomului. Pozitia centromerului este variabila, impartind cromozomul (cromatida) in doua regiuni inegale. In general, centromerii contin heterocromatina constitutiva. Centromerii cromozomilor umani contin secvente lungi de aproximativ 170 nucleotide (ADN satelit a dispuse in tandem si repetate de la 2 000 la 30 000 ori pentru fiecare centromer. ADN centromeric fixeaza proteine specifice, printre care si cele care servesc drept situs de fixare a microtubulilor in procesul de separare a cromozomilor in timpul diviziunii mitotice. Centromerii impart fiecare cromatida in doua cromatide inegale numite brate.
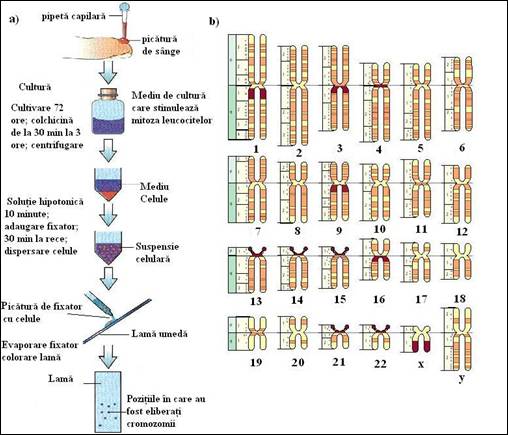
Figura 5. Cariotipul cromozomilor mitotici de la om: a) tehnica folosita pentru obtinerea de preparate de cromozomi mitotici din leucocitele sangelui periferic; b) dupa obtinerea unei fotografii a cromozomilor eliberati din nucleu, acestia sunt aranjati perechi si dupa marime.
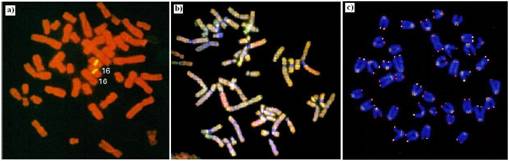
Figura 5. 13. Hibridizarea fluorescenta in situ (FISH) si aplicatiile ei: a) utilizarea de sonde fluorescente pentru marcarea regiunii centromerice a cromozomului 16; b) exemplu de cariotip spectral; c) evidentierea prezentei telomerilor la cromozomi de soarece (adaptat dupa Molecular Cell Biology Lodish H., et. al., 2000)
Telomerii
Cele doua extremitati ale moleculei de ADN, de la nivelul fiecarui cromozom, poseda segmente particulare de secvente repetitive numite telomeri, care formeaza un "coif" la fiecare capat. La om telomerii poseda motivul repetitiv TTAGGG/AATCCC care se repeta de o mie de ori si permite replicarea normala a capetelor cromozomilor (Figura 5.14
Contrar celor mai multe tipuri de secvente repetitive, care variaza mult intre specii, chiar daca ele sunt inrudite si apropiate evolutiv, in cazul telomerilor de la om si de la marea majoritatea a vertebratele studiate pana in prezent, gasim aceeasi secventa telomerica. La alte organisme, cum sunt protozoarele si drojdiile, telomerii au secvente diferite. Totusi, asemeni situatiei de la vertebrate, una dintre catene este intotdeauna bogata in resturi de guanozina iar complementara sa in resturi de citozina. Catena bogata in G este orientata in sensul 5'-3' catre extremitatea cromozomului si depaseste cu 12 la 15 nucleotide extremitatea catenei bogata in C. Datorita existentei acestui dezechilibru catena bogata in G formeaza o coada scurta, monocatenara, la cele doua extremitati ale cromozomului. Aceasta dispozitie persista de la o generatie celulara la alta datorita unei enzime speciale, telomeraza, care poate adauga noi unitati repetitive la extremitatea 3' a catenei bogate in G (Figura 5.14b
Telomeraza, ale carei proprietati au fost bine studiate de catre Elisabeth Blackburn si colegii sai de la Universitatea din California, San Francisco, este o transcriptaza inversa care asambleaza fragmente de ADN folosind o catena ARN matrita. Este o enzima foarte neobisnuita deoarece fragmentul de ARN care serveste drept model (matrita) face parte din structura.
Telomerii au roluri importante: i) sunt necesari replicarii complete a cromozomului; ii) protejeaza cromozomii de nucleaze si de alte influente destabilizatoare; iii) impiedica fuziunea dintre extremitatile cromozomilor in procesele de recombinare; iv) faciliteaza interactiile dintre extremitatile cromozomilor si anvelopa nucleara in anumite tipuri celulare. De asemenea, telomerii si activitatea telomerazei par sa fie unii dintre principalii actori implicati in procesul de imbatranire. Celulele normale, in cultura, nu sunt capabile sa se divida decat de un numar limitat de ori inainte de a da semne de "imbatranire" si de a muri in final. Dintre ipotezele propuse pentru a explica aceasta observatie este si scurtarea progresiva a telomerilor.
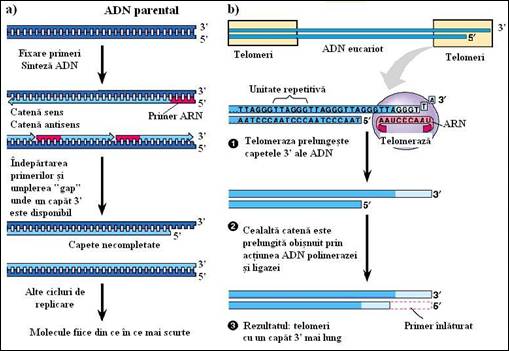
Figura 5. Completarea capetelor ADN de catre telomeraza: a) procesul de replicare si evidentierea formarii capetelor incomplete ale cromozomilor pentru catena intarziata; b) mecanismul de actiune al telomerazei cu evidentierea motivelor repetitive de la capetele cromozomilor
Telomerii se scurteaza deoarece cea mai mare parte a celulelor umane par sa fie lipsite de telomeraze. Contrar celulelor normale, celulele canceroase nu inceteaza sa creasca in cultura si devin "nemuritoare". Unul dintre factorii care ar putea contribui la imortalizarea celulelor maligne este reactivarea telomerazei, care conserva lungimea telomerilor de-a lungul generatiilor. De fapt, cercetarile realizate in acest sens au demonstrat clar ca celulele canceroase poseda o activitate telomerazica care nu poate fi evidentiata in celulele normale. Aceasta descoperire a declansat stimularea cercetarilor privind inhibitorii specifici ai acestei enzime in speranta ca acestia vor avea efect benefic in tratamentul anumitor tipuri de cancer.
Tipuri neobisnuite de cromozomi
Cariogramele de la anumite organisme evidentiaza existenta unor trasaturi neobisnuite care nu sunt intalnite la om. Acestea includ minicromozomii si cromozomii B.
Minicromozomii sunt structuri relativ mici in lungime dar bogate in gene. De exemplu, genomul de la pui contine 39 cromozomi: i) 6 macrocromozomi care contin 66% din cantitatea de ADN si numai 25% dintre gene; ii) 33 minicromozomi care contin 33% ADN si restul de 75% gene. Densitatea genica pe minicromozomi este de sase ori mai mare decat pe macrocromozomi.
Cromozomii B sunt cromozomi aditionali existenti la unii indivizi din populatii, dar nu la toti. Acestia sunt structuri comune la plante dar exista si la fungi, insecte si animale. Cromozomii B par sa fie versiuni fragmentare ale cromozomilor normali care sunt rezultatul unor evenimente neobisnuite aparute in timpul diviziunii nucleare. Unii dintre acestia contin gene, care adesea codifica pentru ARNr, dar nu se cunoaste starea de activare a acestora. Prezenta cromozomilor B poate afecta fenotipul organismului, mai ales la plante, unde sunt asociati cu viabilitatea redusa. O serie de studii au evidentiat pierderea graduala a cromozomilor B in descendenta datorita anormalitatilor evidentiate in modul de mostenire.
Impachetarea genomului eucariot
Dupa cum am evidentiat mai sus, o celula umana normala contine aproximativ 6 miliarde de perechi de baze repartizate intre 46 de cromozomi (cantitatea prezenta in numarul diploid de cromozomi ne-replicati). Fiecare cromozom contine o singura molecula continua de ADN. Cu cat cromozomul este mai mare cu atat este mai lunga molecula de ADN pe care o contine. Deoarece fiecare pereche de baze ocupa aproximativ 0,34 nm, 6 miliarde de perechi de baze corespund unei molecule lungi de 2 metri. De asemenea, in celula, ADN este hidratat cu cantitati importante de apa (aproximativ 6 molecule de apa pentru fiecare pereche de baze), care si acestea duc la cresterea volumului. Problema cheie a fost identificarea modalitatilor prin care este posibila localizarea a 2m de ADN hidratat intr-un nucleu al carui diametru nu depaseste 10 mm, asigurand in acelasi timp, accesul proteinelor si enzimelor implicate in procesele de reglare. O alta problema, la fel de importanta, se refera la modul in care molecula de ADN, dintr-un cromozom, este organizata pentru a nu se innoda cu moleculele altor cromozomi. Raspunsurile la toate aceste probleme il gasim in modul remarcabil de impachetare al moleculei de ADN. Se cunoaste de mult timp ca fibrele care formeaza cromatina, si intra in structura cromozomilor, sunt formate din ADN in asociere cu proteine. Proteinele din structura cromatinei sunt impartite in doua grupe principale: histonele si proteinele cromozomiale nehistonice. Histonele reprezinta o colectie de mici proteine bazice bine definite, in timp ce proteinele nehistonice sunt compuse dintr-un numar mare de proteine diferite cu rol structural, enzimatic si reglator. Cromatina mai contine si un procentaj scazut de ARN compus, in principal, din catene de ARNhn in diferite stadii de maturare si din ARNsn care intervine in procesul de "splicing" al ARNm.
In celulele eucariote, unitatea de baza a organizarii cromatinei este nucleozomul. Acesta reprezinta o entitate in care 146 pb sunt rasucite in doua ture de super-elice stanga in jurul unui octamer de histone care contine cate doua exemplare din moleculele H2A, H2B, H3 si H4. Nucleozomii sunt organizati in filamente de 10 nm in diametru prin interactiuni cu histona H1, care se lega la ADN care intra si iese din nucleozom. Un al treilea nivel de organizare este obtinut prin rasucirea unui filament de 10 nm intr-o elice care contine 6 nucleotide pe tur, cu formarea unui solenoid cu un diametru de 30 nm. Structuri mai complexe ale cromatinei sunt realizate prin condensarea filamentelor de 30 nm, dar detaliile privind aceste structuri sunt mai putin cunoscute (Figura 5.15
Microscopia electronica a furnizat dovezi decisive privind organizarea cromatinei in regiuni, bucle si domenii distincte de 30 la 300 kpb, fixate fiecare pe o matrice bogata in proteine. Fiecare bucla pare sa aiba doar o singura origine de replicare si sa se comporte ca o unitate de replicare. Buclele, sunt unitati de supra-rasucire independente, structura topologica a fiecareia fiind independenta de starea celorlalte. Se pare ca acest lucru este posibil datorita fixarii extremitatilor buclelor in matrice. Cu toate ca fiecare bucla contine mai multe unitati de transcriptie, activitatea intregii regiuni poate fi coordonata pentru a fi reprimata sau potential activata (Figura 5.15
Nucleozomii: primul nivel de organizare al cromozomului
Impachetarea precisa a ADN de la eucariote depinde de histone, grup remarcabil de proteine bazice care se imparte in cinci subgrupe, in principal in functie de continutul lor in lizina si arginina care reprezinta aproximativ ¼ din totalul aminoacizilor (Tabel 5.5). La pH fiziologic, aceste proteine sunt incarcate pozitiv si poseda o structura tertiara cu un grad mare de elice alfa. Aminoacizii bazici din structura sufera modificari post-traductionale (acetilare, metilare, fosforilare) reversibile enzimatic. Studiile comparative de secventa si de structura tridimensionala au condus la divizarea familiei histonelor in doua grupuri: H2A, H2B, H3 si H4 pe de o parte si H1 pe de alta parte. Aceasta eterogenitate este subliniata si de pozitia lor in nucleozom.
De fapt, histonele H4 si H3, si intr-un grad mai mic H2A si H2B, sunt proteine foarte conservate evolutiv. Exemplul cel mai frapant este al histonei H4 de la bovine si de la mazare, care au acelasi numar de aminoacizi (102) si ale caror secvente primare nu difera decat prin doi aminoacizi: o lizina in locul unei arginine si o leucina in locul unei valine. Aceste diferente minore sunt prezente in conditiile in care divergenta dintre animal si planta s-a produs acum un miliard de ani. Structura histonei H1 este mult mai variabila de la o specie la alta. Aceasta conservare extrema sugereaza ca toti aminoacizii au un rol bine definit in functionarea proteinei.
Structura tertiara a histonelor din primul grup demonstreaza existenta a doua domenii: un domeniu central globular, hidrofob si un domeniu N-terminal, hidrofil, incarcat pozitiv si flexibil. Histona H1, mai lunga, poseda un al treilea domeniu la nivelul capatului C-terminal, hidrofil si flexibil.
La inceputul anilor 1970, s-a putut observa ca in urma tratamentului cromatinei cu nucleaze nespecifice, cea mai mare parte a ADN era transformata in fragmente de aproximativ 200 perechi de baze sau in multipli ai acestora.
Din contra, acelasi tratament aplicat moleculelor de ADN "nud" determina obtinerea unei populatii de fragmente de marimi aleatoare. Aceasta descoperire i-a facut pe cercetatori sa considere ca fragmentele uniforme de ADN cromozomial erau protejate de atac enzimatic probabil prin asocierea lor periodica cu o proteina.
Tabel 5. Caracteristicile histonelor din timus de vaca
|
Histone |
Nr. aminoacizi |
Masa kDa |
% Arg |
% Lys |
PUE* (10-6 ani) |
|
H1 | |||||
|
H2A | |||||
|
H2B | |||||
|
H3 | |||||
|
H4 |
* PUE = Perioada unitara de evolutie: durata de timp necesara pentru o schimbare cu 1% a secventei de aminoacizi a unei proteine dupa separarea a doua specii
In 1974, Roger Kornberg, de la Universitatea Harvard, a propus un nou tip de structura secundara pentru cromatina bazandu-se pe rezultatele digestiei cu nucleaze asupra cromatinei din diferite surse. Astazi, se cunoaste ca nucleozomii contin o particula care constituie nucleul nucleozomului, compus din 146 perechi de baze de ADN suprarasucit, care face aproape doua spire in jurul unui miez proteic central care contine 8 molecule din patru histone: H2A, H2B, H3 si H4 (Figura 5. 16d
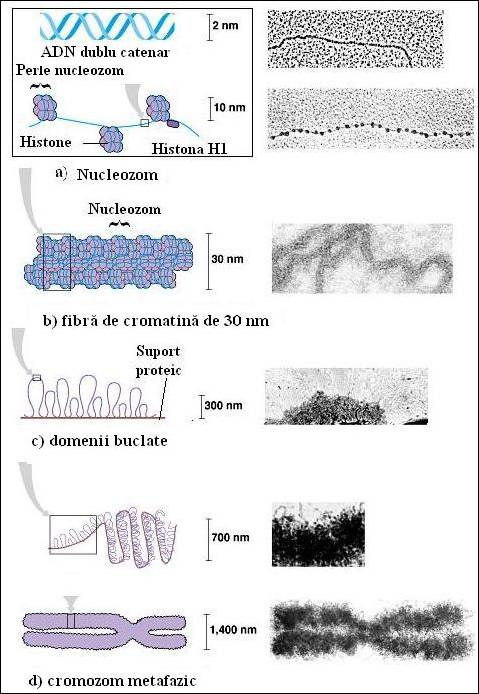
Figura 5. Schema generala a organizarii cromatinei cu evidentierea principalelor nivele cunoscute de organizare (Campbell, et al., Biology 2002)
Particulele de nucleozomi sunt unite unele de altele printr-un segment de ADN de legatura, de lungime variabila, dar care are in general 60 de perechi de baze. ADN din structura unui nucleozom si din bratul de legatura reprezinta aproximativ 200 de perechi de baze, ceea ce corespunde valorii fragmentelor gasite in cursul primelor experiente cu nucleaze (Figura 5.16a si b). Nucleele nucleozomilor, care au un diametru de aproximativ 10 nm, si ADN de legatura, cu un diametru de 2 nm, apar pe micrografiile electronice asemeni "unui sirag de matanii" (Figurile 5.16 si 5.17c). Pentru fiecare nucleozom, o molecula de histona H1 este pozitionata in exteriorul nucleului fiind asociata la cele doua extremitati ale ADN care intra si ies de pe suprafata complexului proteic. (Figura 5.16d). Daca se realizeaza eliminarea selectiva a moleculelor de histone H1, printr-un tratament al fibrelor cu solutii saline de concentratie scazuta, cromatina capata un aspect mult mai dezorganizat.
In general, resturile de aminoacizi bazici ale histonelor nucleului sunt grupate la extremitatile moleculei, restul structurii pastrand un caracter relativ hidrofob. Aceasta separare convine perfect organizarii nucleozomului. Regiunile neincarcate si hidrofobe ale histonelor ocupa centrul particulei si favorizeaza agregarea intr-un nucleu strans. Portiunile polare, bazice, ale moleculelor din nucleu formeaza cozi flexibile orientate catre exteriorul particulei, unde resturile incarcate pozitiv pot stabili interactii ionice cu gruparile fosfat negative din structura scheletului ADN. Cu toate ca, molecula de ADN este strans asociata nucleului format din histone prin suprafata interna a fibrei elicoidale, suprafata sa externa ramane expusa si poate interactiona cu moleculele de reglare.
Studii de cristalografie cu raze X au demonstrat ca resturile incarcate pozitiv sunt reunite sub forma de perechi la suprafata octamerului histonic. Situsurile pereche formeaza un fel de spirala pe drumul pe care se presupune ca il urmeaza elicea ADN care inconjoara nucleul nucleozomului. Acestea servesc drept "puncte de fixare" pentru cele doua catene ale helixului. Deoarece toate moleculele ADN, indiferent de origine si secventa nucleotidica, poseda un schelet riboza-fosfat identic cu cel cu care interactioneaza histonele, nu trebuie sa ne surprinda extrema conservare a moleculelor aminoacizilor din structura histonelor.
Daca pentru spiralizarea a 200 de perechi de baze de ADN sunt necesare aproximativ noua molecule de histone, o celula umana care contine 6 miliarde de perechi de baze de ADN, poate contine aproximativ 300 milioane de histone necesare impachetarii primare a materialului genetic. Astfel se explica, necesitatea unui numar mare de exemplare de gene care codifica pentru histone in celulele care se divid rapid.
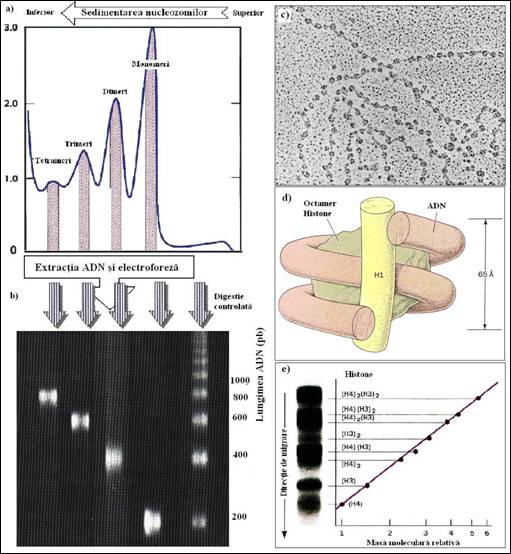
Figura 5. Structura si dimensiunile nucleozomilor: a) sedimentarea nucleozomilor in gradient de sucroza duce la obtinerea unei serii de picuri care corespund formelor monomere, dimere, trimere, etc., ale nucleozomilor; b) ADN extras din fractiile individuale este supus electroforezei impreuna cu un martor ADN digerat cu nucleaze; c) micrografie electronica a cromatinei eliberata dintr-un nucleu al unei celule de Drosophila. Se observa ca fibrele de cromatina sunt formate din nucleozomi conectati intre ei prin segmente scurte de ADN; d) schema care demonstreaza structura unei particule nucleozomale cu o molecula de histona H1 asociata; e) electroforeza in SDS a unui amestec de histone H3 si H4 din timus de vitel. Se observa toate benzile corespunzatoare componentelor tetramerului (H3)2(H4)2 (Lewin B., Genes VII,2000).
Interactia dintre histone si ADN este, in principal, de natura structurala si relativ independenta de secventa nucleotidica. In vitro, un fragment de ADN care in mod normal nu este asociat cu histone, cum este cazul bacteriofagilor sau a polinucleotidelor bicatenare sintetice, formeaza subunitati nucleozomale daca este incubat cu histone purificate care provin de la plante si animale. Experientele de acest tip ilustreaza clar capacitatea de autoasamblare a nucleozomilor.
Chiar daca asamblarea histonelor cu ADN nu depinde de secventa, aceasta nu semnifica neaparat ca pentru o anumita gena, nucleele nucleozomilor sunt localizate la situsuri aleatoare. De fapt, s-a demonstrat ca anumite parti ale genelor manifesta interactiuni constante cu regiunile vecine. Exemplul prezentat in micrografia electronica din figura 5.17 demonstreaza ca pe cromozomul virusului SV40 particulele nucleozomale sunt distribuite periodic, cu exceptia unei regiuni care este lipsita de asocierea cu histonele.
Absenta particulelor nucleozomale, la nivelul acestui fragment, care contine si originea de replicare a cromozomului viral, presupune legarea in acel loc, a unei proteine nehistonice situs-specifica care poate influenta pozitionarea particulelor nucleozomale.
Pozitionarea nucleozomilor poate fi influentata si de capacitatea ADN de a se rasuci in jurul nucleului histonelor. Astfel, segmentele de ADN bogate in A-T sunt mai flexibile si se curbeaza mult mai usor decat regiunile bogate in G-C. In consecinta, nucleozomii au tendinta sa se formeze in regiunile in care raportul A-T si G-C este optim deoarece este diminuata energia necesara pentru rasucirea ADN in jurul octamerilor de histone. S-a constatat ca regiunile bogate in A-T se gasesc preferential in locurile unde fosa mica a dublei elice este orientata catre nucleul histonic, in timp ce regiunile bogate in G-C se gasesc, cu predilectie, in zonele in care fosa mica este orientata catre exterior. De fapt, pozitionarea nucleozomilor poate fi foarte importanta pentru exprimarea genelor.
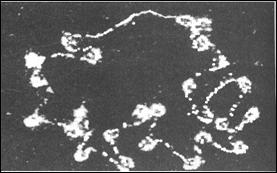
Figura 5. Structura genomului SV40. Se evidentiaza regiunea lipsita de nucleozomi in partea superioara a figurii
O alta problema este modul in care un nucleu cu un diametru de 10 mm poate contine o cantitate de ADN a carei lungime este de 200 000 de ori mai mare decat aceasta valoare. Formarea nucleozomilor reprezinta prima etapa importanta a procesului de condensare. Daca intinderea ar fi completa, cele 200 perechi de baze ale unui nucleozom individual de 10 nm diametru ar ocupa aproximativ de 70 nm, pentru o valoare a planului nucleotidelor de 0,34 nm. In consecinta, raportul ca de condensare a ADN, din structura nucleozomilor este de aproximativ 7:1.
Nivelurile structurale superioare ale cromatinei.
Deci, cel mai redus nivel de organizare al cromatinei este rasucirea moleculei de ADN in jurul inimii nucleozomului si are un de diametru 10 nm. Totusi, cromatina nu se gaseste in celula sub aceasta forma relativ rasucita de "sirag de matanii". Micrografiile electronice ale sectiunilor de nucleu releva un umar de pete minuscule cu un diametru de aproximativ 30 nm care reprezinta sectiuni transversale la nivelul fibrelor de cromatina. Daca se separa cromatina din nucleu, in prezenta ionilor divalenti, se observa prezenta unor filamente de aceeasi grosime (Figura 5.18
Structura acestui filament de 30 nm ramane inca un subiect discutabil. In figura 5.18 este prezentat modelul solenoid de suprarasucire a filamentului subtire al nucleozomilor (10 nm) pentru a forma un filament de ordin superior, mai gros. Se presupune ca, fibrele condensate se formeaza prin impachetarea nucleozomilor intr-o structura spiralata (aranjament solenoid) care contine sase nucleozomi pentru fiecare tur de rasucire. Acest nivel suplimentar de organizare al cromatinei creste de sase ori raportul de condensare. Filamentul de 30 nm este conservat prin interactiunea intre moleculele de histona H1 ale nucleozomilor vecini (Figura 5.18 b
Daca sunt extrase selectiv moleculele H1, filamentele groase de cromatina se deruleaza si se transforma in "margele" (matanii) mai subtiri si mai inguste. Readaugarea histonei H1 restabileste structura de ordin superior. Studii recente de microscopie electronica sugereaza ca fibrele de 30 nm sunt mai putin uniforme decat modelul solenoid imaginat. Cromatina condensata poate fi de fapt foarte dinamica fiind formata din structuri partial depliate care se pot replia rapid in structuri solenoide ocazionale.
Cromatina din regiunile cromozomiale care nu sunt transcrise frecvent este predominant in forma condensata de 30 nm, in timp ce regiunile transcrise activ se considera ca adopta forma relaxata de "sirag de matanii" (Figura 5.18 a
Urmatoarea etapa de condensare a ADN in nucleu il reprezinta organizarea filamentelor de 30 nm intr-o serie de bucle mari suprarasucite. Se estimeaza ca fiecare bucla contine intre 10 si 150 kilobaze ADN. Se pare ca acestea sunt fixate la baza cu proteine specifice printre care si o topoizomeraza de tip II care intervine pentru a controla gradul de spiralizare al ADN la nivelul buclei. Topoizomeraza elibereaza moleculele de ADN daca o bucla se incurca. Buclele de ADN ar putea imparti genomul in "domenii", fiecare continand un lot mic de gene care sunt exprimate printr-un mecanism comun de reglare. Normal, buclele de cromatina sunt etalate la interiorul nucleului si nu sunt vizibile, dar prezenta lor poate fi evidentiata in anumite circumstante. De exemplu, daca tratam cromozomii mitotici izolati cu reactivi care extrag histonele, putem observa ca ADN eliberat de histone se pliaza sub forma de bucle plecand de la esafodaj proteic, format din proteine nehistonice (Figura 5.15
Micrografia electronica din figura 5.19c evidentiaza buclele lungi de ADN ancorate la un schelet cromozomial compus din proteine nehistonice obtinut prin indepartarea histonelor din structura cromozomilor din celule HeLa, in metafaza. Acest esafodaj flexibil are forma cromozomului metafazic si persista chiar si atunci cand ADN este digerat cu nucleaze (Figura 5.19c
Acelasi tip de bucle se pot evidentia in cromozomii politeni interfazici din celulele insectelor si in cromozomii meiotici "in perie" ("lamp-brush") din ovocitele de amfibieni (Figura 5.19a), ceea ce demonstreaza ca acestea nu sunt proprii cromozomilor mitotici.
O serie de experimente de hibridizare "in situ", realizate pe celule umane in interfaza, utilizand diferite sonde fluorescente, au dus la imaginarea modelului prezentat in figura 5.19b. Astfel, s-a evidentiat ca o serie de sonde (A la H) care recunosteau secvente situate la milioane de perechi de baze distanta, in structura ADN linear, erau pozitionate foarte aproape unele de altele in structura cromozomului interfazic. Se considera ca apropierea dintre aceste situsuri, numite si "regiuni asociate scheletului" (denumite SARs de la "scaffold-associated regions" sau MARs de la "matrix-attachment regions") este posibila datorita asocierii lor la scheletul cromozomului. In general, regiunile asociate scheletului se gasesc intre unitatile de transcriptie. Deci, genele sunt localizate prioritar la nivelul buclelor de cromatina ale caror baze sunt atasate scheletului cromozomului.
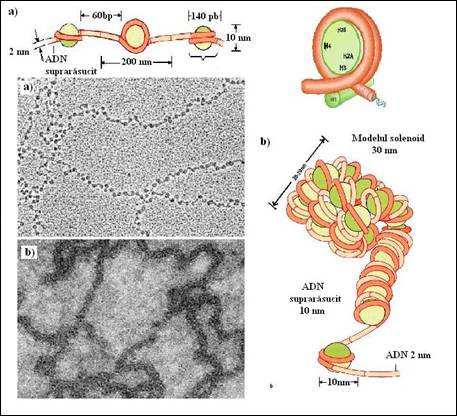
Figura 5.18. Modelul solenoid de organizare a fibrelor de cromatina condensata de 30 nm: a) micrografia si modul de organizare a nucleozomilor din structura; b) micrografie prezentand fibrele de 30 nm si modelul de impachetare solenoid cu caracteristicile sale structurale (adaptat dupa Molecular Cell Biology Lodish H., et. al., 2000)
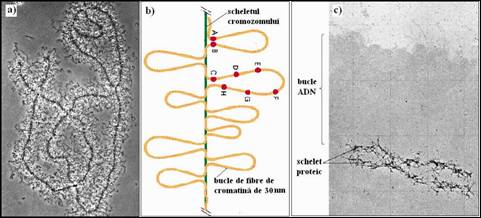
Figura 5. Nivelurile structurale superioare ale cromatinei: a) micrografie electronica prezentand domenii de cromatina, in bucle la cromozomii in perie izolati dintr-un ovocit de amfibian; b) model de structurare a buclelor fibrelor de cromatina de 30 nm imaginat in urma experientelor de hibridare in situ cu sonde (A la H) situate la distante variabile pe ADN liniar; c) micrografie electronica a unui cromozom metafazic din celule HeLa, lipsit de histone ca urmare a unui tratament moderat cu detergenti.
Cromozomul mitotic reprezinta etapa ultima de condensarea a cromatinei (Figura 5.11 si 5.17). De obicei, un mm de cromozom mitotic contine un cm de ADN. Aceasta condensare, realizata ca urmare a unui proces inca putin cunoscut, este insotita de fosforilarea practic a tuturor moleculelor de histone H1 la nivelul celor cinci resturi de serina din molecula.
O imagine generala a diferitelor niveluri de organizare ale cromatinei de la filamentele nucleozomale pana la cromozomii mitotici este prezentata in figura 5.17.
Heterocromatina si eucromatina
Dupa terminarea mitozei, cea mai mare parte a cromatinei care compune cromozomii mitotici foarte condensati se disperseaza si revine la starea sa interfazica difuza. Totusi, in cea mai mare parte a celulelor, aproximativ 10% din materialul cromozomial isi pastreaza forma condensata compacta pe tot parcursul interfazei, cromatina condensata putand fi evidentiata la periferia nucleului. Termenul de heterocromatina desemneaza cromatina care ramane condensata in timpul interfazei pentru a o distinge de eucromatina care desemneaza starea dispersata. Daca celulele sunt puse in prezenta unui precursor ARN radiomarcat, la nivelul uridinei (uridina 3H), si apoi fixate, sectionate si observate dupa autoradiografie, se constata ca regiunile de heterocromatina raman nemarcate deoarece acestea sunt putin transcrise sau chiar netranscrise. Cercetarile din ultimii ani demonstreaza clar posibilitatea trecerii reversibile dintr-o forma in alta, aceasta clasificare fiind mai degraba didactica.
Numeroase gene de la eucariotele multicelulare nu sunt exprimate decat in anumite momente ale dezvoltarii si/sau in anumite tesuturi. Deci, celulele trebuie sa posede modalitati eficace pentru a reprima expresia tuturor genelor care nu sunt caracteristice unui anumit tip de celula intr-o anumita etapa a dezvoltarii sale. Represia unor blocuri mari de gene, prin limitarea disponibilitatii factoriilor de transcriptie, este un mecanism destul de putin probabil pentru o astfel de reglare. Ideea este sustinuta de faptul ca unele gene, care nu sunt exprimate in mod normal intr-o celula, pot fi transcrise daca sunt introduse prin transfectie in aceasta. De exemplu, genele globinei, induse prin transfectie in fibroblaste, sunt exprimate de o mie de ori mai puternic decat genele rezidente ale globinei. Aceasta diferenta persista in cursul diviziunilor succesive daca genele transfectate sunt integrate la nivelul unor situsuri cromozomiale. De fapt, posibilitatea ca genele transfectate sa aiba o activitate mai mare decat genele endogene este un efect destul de obisnuit dar nu universal. Este general admis ca aceasta represie este rezultatul sechestrarii genelor "silentioase" in complexe de cromatina superior structurate care le fac inaccesibile masinariei de transcriptie.
In absenta informatiei privind organizarea acestei structuri a cromatinei, este imposibil de descris starea de tranzitie care separa cromatinele reprimate si exprimate. O indicatie este furnizata de corelatia care exista intre momentul in care gena este replicata si cel in care ea poate fi transcrisa. Genele de mentinere, exprimate in permanenta in toate celulele (exemplu: dihidrofolat-reductaza, actina citoplasmatica, glucozo-6-fosfat dehidrogenaza) sunt replicate in prima jumatate a fazei S. In acelasi context, genele neexprimate tind sa fie replicate mai tarziu. De asemenea, replicarea genelor este precoce in tesuturile in care ele sunt exprimate si tarzie in tesuturile in care acestea sunt "silentioase'. DE exemplu, la mamifere cromozomul X inactiv este replicat dupa cromozomul X activ. Aceste corelatii sugereaza ca replicarea precoce creeaza o stare mai accesibila masinariei de transcriptie crescand eficienta fixarii factorilor de transcriptie.
Heterocromatina
Heterocromatina poate fi divizata in doua categorii: constitutiva si facultativa, in functie de permanenta starii condensate. Heterocromatina constitutiva ramane condensata tot timpul si reprezinta regiunile din structura ADN care raman silentioase permanent. In celulele de mamifere, cea mai mare parte a heterocromatinei constitutive se gaseste in apropierea centromerului tuturor cromozomilor si in alte cateva regiuni particulare, cum este bratul distal al cromozomului Y la masculi. La multe plante, extremitatile cromozomului (telomerii) sunt si ele formate din heterocromatina constitutiva. In principal, ADN din structura heterocromatinei constitutive este format din secvente inalt repetitive si se presupune ca este lipsit de gene care codifica pentru proteine. Se considera ca, heterocromatina contine elemente a caror influenta se face simtita la o anumita distanta si poate afecta starea fiziologica a genelor invecinate. Astfel, daca unele gene active, care si-au schimbat pozitia ca urmare a unei transpozitii sau a unei translocari, se afla pozitionate in apropierea heterocromatinei, ele prezinta tendinta de a deveni inactive. Acest fenomen poarta numele de efect de pozitionare.
Contrar formei constitutive, heterocromatina facultativa este inactivata specific in cursul anumitor stadii de viata ale organismului. Un exemplu tipic de heterocromatina facultativa este furnizat de inactivarea unui cromozom X in celulele femele de la mamifere. Celulele masculine au un mic cromozom Y si un cromozom X mult mai mare. Cromozomii X si Y au putine gene in comun astfel incat barbatii nu poseda decat un exemplar al genelor situate pe cromozomii sexuali. Desi, celulele de la femele contin doi cromozomi X, unul singur este functional in procesul de transcriptie. Celalalt cromozom ramane condensat sub forma unei mase de heterocromatina (Figura 5.20b) numit corpuscul Barr, dupa numele cercetatorului care l-a descoperit in anul 1949. Se considera ca, producerea corpusculului Barr este un mecanism normal, gratie caruia celulele femele si mascule dispun de acelasi numar de cromozomi X activi si sintetizeaza cantitati echivalente de produsi codificati de genele situate pe cromozomul X.
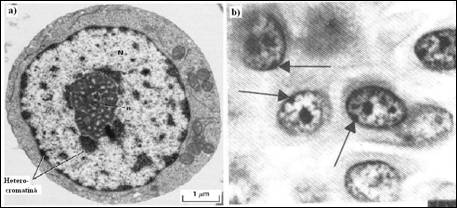
Figura 5. . Evidentierea heterocromatinei: a) micrografie electronica a unei celule stem din maduva spinarii. Regiunile inchise de la periferia nucleului si din afara nucleolului reprezinta heterocromatina; b) prezenta cromozomului X inactivat sub forma corpusculului Barr intr-o celula de femeie.
Inactivarea cromozomului X
In 1961, Mary Lyon emite ipoteza ca un cromozom X devine heterocromatic in toate celulele femelelor de mamifere intr-un stadiu precoce al dezvoltarii embrionare. Acest mecanism a fost numit inactivarea cromozomului X si s-a demonstrat ulterior ca se produce in embrioni, in momentul gastrulatiei. Deoarece, inactivarea in embrion este un proces aleator, atat cromozomul X de origine materna cat si cel de origine paterna prezinta aceeasi probabilitate de inactivare in oricare dintre celule. In consecinta, cromozomul X patern poate fi inactivat intr-o celula embrionara in timp ce cromozomul X matern poate fi inactivat intr-o celula vecina. Din acel moment, acelasi cromozom X este inactiv in toata descendenta unei celule particulare. Cel de al doilea cromozom X este reactivat, in celulele germinale, inainte de inceputul meiozei. Deci, cei doi cromozomi X sunt activi in timpul ovogenezei si toti gametii primesc un cromozom X activ.
Deoarece, cromozomii X care provin de la tata si de la mama pot purta alele diferite pentru acelasi caracter, femelele adulte sunt, intr-un anumit sens, un mozaic genetic, alele diferite putand functiona in celule diferite. Mozaicismul cromozomului X se traduce prin pete de culoare in blana anumitor mamifere, cum sunt pisicile "calico". Din fericire, la om, genele de pigmentare nu sunt localizate pe cromozomul X, de unde absenta "femeilor calico". Existenta mozaicismului datorat inactivarii cromozomului X a fost demonstrata in cazul ochilor unor femei heterozigote pentru daltonism rosu-verde. Daca ochii sunt luminati de un fascicol de lumina rosie sau verde, se pot detecta in retina, regiuni de celule care prezinta o proasta perceptie a culorilor, dispersate printre celule unde vederea este normala.
Mecanismul responsabil de inactivarea cromozomului X a atras atentia dupa ce in 1992 o publicatie a sugerat ca inactivarea este initiata de o molecula de ARN transcrisa plecand de la gena Xist localizata pe cromozomul X inactiv si nu de proteina codificata de aceasta. De asemenea, s-a evidentiat ca gena omologa situata pe cromozomului X activ nu este transcrisa. Acesta reprezinta un exemplu rar de expresie genica dintr-o regiune cromozomiala inactiva (heterocromatina). Aceasta descoperire, si multe altele, au stat la originea ipotezei conform careia moleculele de ARN pot functiona ca reglatori directi ai genelor, ceea ce reprezenta o functie inca necunoscuta a ARN.
Sensibilitatea la nucleaze
Judecand dupa sensibilitatea la nucleaze (deoxiribonucleaza I sau nucleaza din Micrococus), regiunile cromatinei active transcriptional sunt mai putin compacte decat cele ale genelor silentioase (inactive). Aceasta sensibilitate la nucleaze este adesea caracteristica pentru regiuni care cuprind mai multe mii de perechi de baze de o parte si de alta a unitatii de transcriptie. Nu au fost total identificate bazele moleculare ale acestei sensibilitati diferite. Studiile biochimice sugereaza ca nucleozomii genelor active sunt saraci in histone H1 si bogati in histone acetilate, legate prin ubiquitina si complexate cu mai multe tipuri de proteine HMG acide (HMG pentru High Mobility Group). Aceste modificari altereaza asocierea histonelor cu ADN si ar putea favoriza separarea neregulata a nucleozomilor, facand ADN mai sensibil la digestie. De exemplu, periodicitatea normala a situsurilor nucleazice de clivare ale genelor proteinelor de soc termic este modificata in timpul inducerii expresiei acestora prin cresterea temperaturii. Aceeasi variatie a situsurilor de clivare se poate observa pentru gena ovalbuminei, in oviduct si in ficat, si pentru genele IgL (k) din limfocite, dupa reorganizare intr-o forma transcriptionala activa.
Hipersensibilitatea la nucleaze
Genele transcrise activ prezinta pe langa sensibilitatea normala la nucleaze, situsuri de hipersensibilitate situate in jurul unitatilor de transcriptie. O astfel de hiper-sensibilitate la deoxiribonucleaza I nu apare la nivelul unor fragmente de ADN "nud". In general, situsurile de hipersensibilitate sunt pozitionate imediat in amonte de situsul de initiere a transcriptiei, dar si la extremitatea 3' a unitatii de transcriptie. Acestea pot sa contina secvente de amplificare sau de terminare. In cea mai mare parte a cazurilor, hipersensibilitatea este restransa la tesuturile sau celulele in care genele sunt exprimate sau programate sa fie exprimate. De exemplu, situsul de hipersensibilitate situat in 5' de gena pre-proinsulinei, este prezent in insulele b ale celulelor pancreatice dar nu a fost identificat in celulele hepatice. La fel, au fost evidentiate situsuri de hipersensibilitate asociate genelor reorganizate ale imunoglobulinelor in celulele B, dar nu si in tesuturile hematopoietice.
Probabil, situsurile de hipersensibilitate la nucleaze apar acolo unde exista o discontinuitate in aranjarea nucleozomilor. Pentru prima data, acest fapt a fost demonstrat de corelatiile evidentiate intre situsurile de hipersensibilitate si zona lipsita de nucleozomi din genomul SV40 (Figura 5.17). Aceasta regiune se intinde de la situsul de initiere a transcriptiei genei precoce pana la extremitatea din amonte a elementului activator. Daca acest segment de 400 pb este transferat intr-o alta regiune a genomului SV40 sau este integrat intr-o plasmida, se formeaza in aceste locuri, o zona libera de nucleozomi, hipersensibila la deoxiribonucleaza I. Deci, cele doua proprietati sunt probabil consecinta secventei nucleotidice a acestui segment de ADN. Totusi, este posibil ca sensibilitatea la nucleaza sa fie modulata si de proteine nehistonice care recunosc aceasta regiune.
De asemenea, situsuri de hipersensibilitate la nucleaze si lipsite de nucleozomi au fost identificate in structura elementelor activatoare ale genelor pentru IgH si IgL(k) si in secventele activatoare din amonte asociate extremitatii 5' a genelor metalotioneinei. Alte situsuri de hipersensibilitate la deoxiribonucleaza I au fost asociate cu activarea transcriptiei ADNr. Ele au fost identificate in amonte de situsul de initiere si in apropierea regiunilor separatorilor intergenici. Astfel, hipersensibilitatea la nucleaze a devenit un indicator al regiunilor de reglare a transcriptiei genelor eucariote.
Deoarece localizarea secventelor care regleaza transcriptia coincide cu situsurile de hipersensibilitate la nucleaze si cu situsurile lipsite de nucleozomi, se considera ca aceste caracteristici sunt corelate. Este probabil, ca fixarea factorilor de transcriptie sa deplaseze nucleozomii sau sa altereze impachetarea lor avand drept consecinta crearea unui "vid" identificabil in aranjarea cromatinei sau chiar perturbari mai subtile care sa permita actiunea nucleazelor. Daca impachetarea densa a nucleozomilor este stabilizata de catre histona H1, sau prin replierea sub forma unei structuri mai elaborate, accesul la secventele semnal al proteinelor esentiale transcriptiei poate fi restrans, impiedicand astfel initierea transcriptiei.
In acest domeniu al cercetarilor privind diferentele dintre cromatina exprimata si cea reprimata, precum si a trecerii de la o stare la alta se ridica o serie intreaga de intrebari care, pentru moment, nu si-au gasit raspuns. Pana in prezent stim ca exista regiuni ale cromatinei care pot fi mentinute in stare condensata si neexprimata timp de mai multe cicluri de replicare dupa care sunt disociate si devin active sub actiunea unor stimuli exogeni sau endogeni adecvati. Pentru moment nu putem afirma ca activarea regiunilor specifice ale cromatinei implica interventia unor factori (proteine) care actioneaza pozitiv, sau eliminarea altora care au rol represor, sau ambele situatii.
Aberatiile cromozomiale
O serie de mutatii accidentale care modifica informatia continuta in genele individuale apar cel mai adesea in cursul replicarii ADN sau datorita actiunii unor agenti de mediu. Fara a minimaliza importanta acestora, pentru buna functionare a organismului, trebuie sa aratam ca modificari mult mai importante pot suporta cromozomii, cel mai adesea in timpul diviziunilor celulare. Astfel, bucati de cromozomi se pot pierde sau segmente importante se pot schimba intre cromozomi diferiti. Aceste aberatii cromozomiale apar ca urmare a unor rupturi iar frecventa lor este crescuta in urma expunerii la agenti de mediu capabili sa modifice ADN: infectiile virale, radiatiile X, agentii chimici, etc. De asemenea, cromozomii unor indivizi prezinta zone "fragile" care au o susceptibilitate particulara la rupere. Anumite maladii ereditare rare, ca sindromul Bloom, anemia Fanconi, si ataxia telangiectazica induc o instabilitate a cromozomilor care creste mult aceasta instabilitate.
Consecinta unei aberatii cromozomiale depinde de genele afectate si de tipul de celula atinsa. Daca modificarea intervine intr-o celula somatica ne-reproducatoare, consecintele sunt in general minore, deoarece numai cateva celule ale organismului vor fi afectate. Totusi, in cazuri rare, o celula purtatoare de aberatii se poate transforma intr-o celula maligna si poate dezvolta o tumora canceroasa.
Pot fi transmise generatiei urmatoare, aberatiile cromozomiale care apar in timpul meiozei, particular dupa un crossing-over anormal. Daca un cromozom anormal este transmis printr-un gamet, toate celulele descendentei sale vor avea aberatia si, in general, individul nu depaseste stadiul dezvoltarii embrionare.
Exista mai multe tipuri de anomalii cromozomiale:
Inversiuni: un cromozom este adesea rupt in doua locuri si segmentul situat intre cele doua pozitii este reatasat in ordine inversa. Pana la 1% din oameni sunt purtatori ai unei inversii care poate fi detectata in urma realizarii unui cariotip. Inversiunile pot fi de doua tipuri: i) paracentrica, daca implica numai un brat al cromozomului; ii) pericentrica, daca rupturile se produc de o parte si de alta centromerului (Figura 5.21). Deoarece, de obicei, un cromozom care contine o inversie poseda toate genele cromozomului normal, individul nu este intotdeauna afectat defavorabil. Totusi, daca o celula cu o inversie cromozomiala intra in meioza, cromozomul aberant nu se poate imperechea corect cu omologul sau normal deoarece ordinea genelor este diferita. In acest caz, imperecherea cromozomiala este insotita de formarea unei bucle. Daca in bucla se produce un crossing-over, gametii rezultati din meioza poseda o copie suplimentara a anumitor gene si apare o duplicare. Daca un gamet care contine un cromozom modificat fuzioneaza cu un gamet normal in momentul fecundarii, zigotul rezultat poarta un dezechilibru la nivelul numarului de cromozomi, care de obicei este invariabil.
Translocari: atasarea unui cromozom sau a unui fragment de cromozom la un altul (Figura 5.21). Ca si inversiunile, translocarile care survin intr-o celula somatica pot sa nu aiba deloc efect asupra functionarii celulei si a descendentei sale. Totusi, anumite evenimente de acest tip cresc probabilitatea ca celulele sa devina canceroase. Exemplul cel mai bine cunoscut este cel al cromozomului Philadelphia, pe care il gasim in celulele maligne (nu si in celulele normale) ale indivizilor care sufera de anumite forme de leucemie. Cromozomul Philadelphia, denumit astfel deoarece a fost descoperit in acest oras in 1960, reprezinta o versiune scurtata a cromozomului 22 de la om. De-a lungul anilor, s-a crezut ca segmentul care lipsea reprezenta o simpla deletie dar, odata cu ameliorarea tehnicilor de observare a cromozomilor, s-a demonstrat ca fragmentul care lipsea era transferat unui alt cromozom (nr. 9). Cromozomul 9 poseda o gena care codifica pentru o protein-kinaza care intervine in proliferarea celulara. Ca urmare a translocarii, o mica parte a acestei proteine este inlocuita prin aproximativ 600 aminoacizi suplimentari codificati de catre fragmentul translocat care provine de pe cromozomul 22. Aceasta noua proteina, mult mai lunga, conserva activitatea catalitica a formei originale, dar nu mai este supusa mecanismelor de reglare normala ale celulei (Figura 5.22
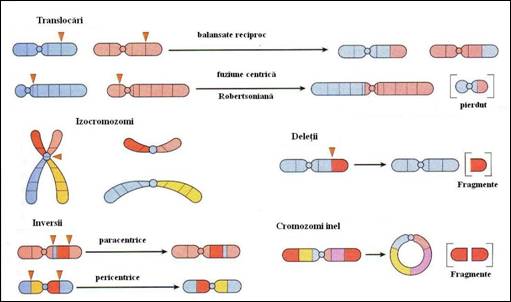
Figura 5.21. Tipuri de rearanjamente cromozomiale
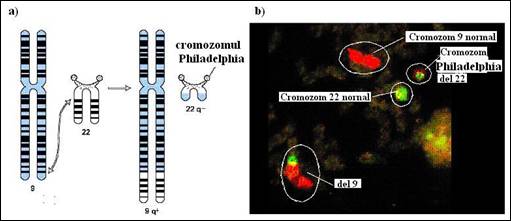
Figura 5.22. Analiza translocarilor cromozomiale prin bandare (a) si prin marcarea fluorescenta a cromozomilor (b). Se observa formarea cromozomului Filadelfia datorita translocarii unei regiuni din cromozomul 22 pe cromozomul 9.
Ca si inversiunile, translocarile sunt o sursa de probleme in procesul de meioza. Continutul genetic al unui cromozom alterat prin translocare este diferit de cel al omologului sau. In consecinta, gametii formati in urma meiozei poseda copii suplimentare ale anumitor gene sau sunt lipsite de acestea. In figura 5.21 sunt prezentate doua tipuri de translocari: i) translocare reciproca balansata, in care se produce cate o singura ruptura pe cei doi cromozomi, cu schimb de material genetic. O astfel de translocare nu poate fi evidentiata fara utilizarea unor tehnici de bandare avansate; ii) fuziune centrica sau translocare robertsoniana, care presupune fuziunea dintre doi cromozomi acrocentrici. De obicei, ruperile apar in apropierea centromerilor fiecarui cromozom. Transferul de segmente duce la formarea unui cromozom foarte mare si a unuia foarte mic care se pierde.
S-a demonstrat ca translocarile joaca un rol important in evolutie producand modificari de mare anvergura care pot fi la originea separarii liniilor evolutive distincte provenite dintr-un stramos comun. Acest accident genetic s-a produs probabil in timpul propriei noastre istorii evolutive recente. Comparatia celor 23 perechi de cromozomi ale celulelor somatice de la om cu cele 24 perechi de cromozomi de la cimpanzeu, gorila si urangutan evidentiaza asemanari frapante. Observarea precisa a doi cromozomi de maimuta, care nu au echivalenta la om, demonstreaza ca in ansamblu acestia corespund, banda cu banda, cromozomului 2 de la om. Se presupune ca, intr-un anumit moment al evolutiei, un cromozom intreg a fost translocat catre altul si a dat nastere unui singur cromozom prin reducerea numarului haploid de la 24 la 23.
Deletiile: reprezinta pierderea unui fragment de cromozom. In functie de localizare acestea pot fi: i) terminale care sunt rezultatul unei singure rupturi la nivelul unui brat al cromozomului, producand un fragment fara centromer care este inlaturat; ii) interstitiale care sunt rezultatul a doua ruperi si care se soldeaza cu pierderea regiunii dintre acestea (Figura 5.21
Adesea, zigotii cu o deletie cromozomiala se formeaza daca un gamet provine dintr-o meioza anormala. De obicei, lipsa unei portiuni cromozomiale antreneaza pierderea de gene esentiale si are consecinte severe, chiar daca cromozomul omolog este normal. Cea mai mare parte din embrionii umani, purtatori ai unei deletii semnificative, nu ajung la termen iar cei care reusesc prezinta adesea numeroase malformatii. In 1963, Jerome Lejeune (descoperitorul sindromului Down) a fost primul care a stabilit o relatie intre o malformatie la om si o deletie cromozomiala. Lejeune a constatat ca un copil cu diferite malformatii ale fetei si o dezvoltare anormala a laringelui era lipsit de o parte a cromozomului 5. Boala a fost denumita "tipatul pisicii" deoarece existenta unei deficiente a laringelui facea ca tipatul copilului sa seamene cu cel al unei pisici care sufera.
Cromozomul sub forma de inel reprezinta o forma speciala de deletie. Aceasta anomalie apare cand la ambele capete ale cromozomului se produc rupturi urmate de fuziunea regiunilor afectate (Figura 5.21). Daca se pierd fragmente importante apar anomalii fenotipice. Din pacate, cromozomii sub forma de inel nu se normalizeaza nici in meioza si nici in mitoza, avand drept rezultat consecinte serioase asupra organismului.
Duplicarile: presupun repetarea unei portiuni de cromozom. Se discuta problema rolului acestora in formarea familiilor multi-genice. Unele duplicari cromozomiale, mai importante, sunt responsabile de prezenta mai multor exemplare ale anumitor gene (ex, trisomie partiala). Cele mai cunoscute duplicari afecteaza genele globinei. Deoarece echilibrul activitatilor metabolice ale organismului este foarte sensibil, existenta unor exemplare suplimentare pentru anumite gene poate avea consecinte negative grave.
Izocromozomii: se obtin cand unul dintre bratele cromozomului este pierdut si bratul care ramane este duplicat. Se obtine un cromozom care contine doua brate scurte sau doua lungi, identice (Figura 5.21). Cele mai frecvente cazuri se intalnesc pentru cromozomul X.
Marimea genoamelor nucleare eucariote
Genoamele nucleare eucariote au marimi variabile (Tabel 5. 1) care pot varia de la 10 Mpb la peste 100 000 Mpb. Adesea, marimea acestora coincide cu complexitatea organismelor, genoamele organismelor eucariote fiind mai mari decat cele ale eucariotelor inferioare. Totusi, marimea nu este determinata numai de numarul de gene ci si de cantitatea de secvente de ADN repetitiv. Cele mai mari genoame au un numar de copii de secvente repetitive este foarte mare.
Dupa cum am constatat in paragrafele anterioare, la eucariote genomul nuclear este impartit intr-un set de molecule de ADN lineare, fiecare fiind impachetata in structura unui cromozom. Din acest punct de vedere nu au fost evidentiate exceptii in lumea eucariotelor, toate organismele cunoscute avand cel putin doi cromozomi in care molecula de ADN este liniara. Variabilitatea la acest nivel se manifesta numai in ceea ce priveste numarul de cromozomi, care pare sa nu fie corelat cu trasaturile biologice ale organismului (Tabel 5.6
Tabel 5. Numarul de cromozomi la diferite organisme (celula haploida)
|
Organism |
Marime genom (Mpb) |
Numar. cromozomi |
|
Saccharomyces cerevisiae | ||
|
Drosophila melanogaster | ||
|
Om | ||
|
Porumb | ||
|
Salamandra |
De exemplu, drojdia Saccharomyces cerevisiae, un eucariot relativ simplu are de patru ori mai multi cromozomi decat musculita Drosophila melanogaster. De asemenea, numarul cromozomilor nu pare sa fie corelat cu marimea genomului sau cu pozitia evolutiva a organismului. Anumite salamandre poseda genoame de aproximativ 30 de ori mai mari decat omul dar au un numar de cromozomi redus la jumatate. O situatie similara se poate consta si in cazul unor plante, cum este porumbul (Figura 5.23 si Tabelul 5.6). Aceste comparatii sunt foarte interesante, dar din pacate nu sunt destul de informative pentru genoamele in sine. Adesea, ele reprezinta numai o reflectare a neuniformitatii evenimentelor evolutive care au modificat arhitectura genomica la diferite organisme.
Complexitatea morfologica a ADN se poate corela cu valoarea C, care reprezinta cantitatea de ADN din genomul haploid. Pe de alta parte complexitatea morfologica a unui organism ar trebui sa reflecte complexitatea genetica. Cu toate acestea, corespondenta dintre cei doi parametrii este cunoscuta drept "Paradoxul Valorii C", deoarece unele organisme au valori C neasteptat de mari. De exemplu genomul pestelui-plaman este de 10-15 ori mai mare decat cel de la mamifere. Adesea paradoxul valorii C este aplicabil si unor specii inrudite (de exemplu valorile C de la diferite specii de drosofile sunt uneori foarte diferite).
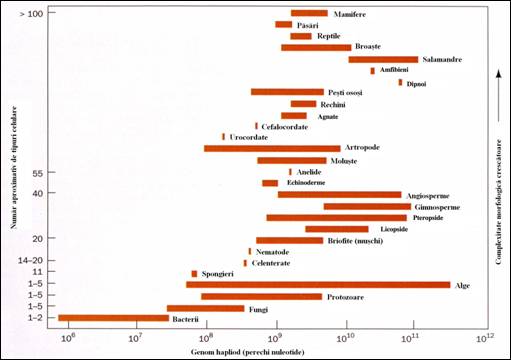
Figura 5.23. Marimea continutului in ADN a genomului haploid de la diferite organisme. Complexitatea morfologica a organismelor, estimata in functie de numarul tipurilor celulare pe care le prezinta, creste de jos in sus (dupa Molecular Cell Biology Lodish H., et. al., 2000)
Analiza moleculara incearca sa explice si paradoxul aparent pe care il reprezinta faptul ca genoamele eucariotelor par sa contina o cantitate insemnata de ADN care nu este necesar celulei. Acum mai bine de 30 de ani se considera ca eucariotele contin aproximativ 50 000 gene. Cum fiecare secventa codanta era considerata ca avand, in medie, 1 200 pb, genomul acestora ar fi trebuit sa contina numai aproximativ 6 x 107 pb. Acum se cunoaste ca genoamele unor ciuperci sunt mai mici decat aceasta valoare si ca cea mai mare parte a genoamelor de mamifere si de plante contin de 50 la 100 ori mai mult ADN (Tabel 5.1
Initial, pentru estimarea numarului de gene, o serie de cercetatori au extrapolat situatia cunoscuta de la E. coli unde de 4,7 milioane pb codifica pentru 3000 de gene. Pastrand proportionalitatea ei considerau ca genomul haploid uman de 2,9 miliarde pb trebuia sa aiba un numar mult mai mare de gene (de 600 ori mai mare). Odata cu incheierea secventializarii genomului uman a fost posibila identificarea tuturor fazelor de lectura si realizarea unei estimari cat mai corecte a numarului genelor de la om. In prezent se discuta de 30-40 000 in loc de 60-80 000 gene cum se considera in momentul inceperii acestui program. De asemenea, evolutia tehnicilor de biologie moleculara si utilizarea unor aparate de secventializare performante permite definitivarea secventializarii unui numar din ce in ce mai mare de organisme si estimarea din ce in ce mai corecta a numarului genelor continute de acestea. In tabelul 5. 7 se realizeaza o prezentare marimii unora dintre cele mai cunoscute genoame, a numarului genelor precum si a distributiei acestora/Mb.
Tabel 5. Marimea genoamelor, numarul de gene si distributia acestora
|
Organism |
Marime genom |
Nr. de gene estimat |
Nr. de gene/Mb |
|
H. influenzae (bacterie) |
1,8 Mb | ||
|
S. cerevisiae (drojdie) |
12 Mb | ||
|
C. elegans (nematod) |
97 Mb | ||
|
A. thaliana (planta) |
100 Mb | ||
|
D. melanogaster (musculita otet) |
180 MB | ||
|
Homo sapiens (om) |
3 200 Mb |
Aceasta variatie inexplicabila a marimii genoamelor apare deoarece cromozomii eucariotelor contin, pe langa informatia genetica din structura genelor, cantitati variabile de ADN, plasate in: i) introni; ii) in regiunile intra-genice si inter-genice; iii) regiuni repetitive in tandem sau dispersate pe toata lungimea genomului. Acest ADN, a carui functie nu a fost inca demonstrata, este adesea compus din repetitii de secvente, unele dintre ele nefiind niciodata transcrise. El mai este denumit si " ADN nefunctional". In tabelul 5.8 sunt trecute in revista principalele tipuri de secvente prezente in structura genomului eucariotelor.
Tabel 5. Clasificarea secventelor ADN de la eucariote
|
Gene care codifica pentru proteine Gene solitare Gene duplicate si divergente (familii de gene si pseudogene nefunctionale) |
|
Gene repetitive in tandem care codifica pentru ARNr, ARNr 5S, ARNt si histone |
|
Repetitii ADN Secvente simple ADN ADN moderat repetitiv (elemente de ADN mobile) Transpozomi Retrotranspozomi virali Secvente repetitive lungi dispersate (LINES de la "Long Interspersed Elements; retrotranspozomi nevirali) Secvente repetitive scurte dispersate (SINES de la "Short Interspersed Elements; retrotranspozomi nevirali) |
|
Alte tipuri de secvente separatoare intergenice |
Marimea, adesea excesiva a genoamelor eucariote se poate datora urmatoarelor aspecte: i) cea mai mare parte a genelor care codifica pentru polipeptide poseda cel putin un intron, un numar mare de gene avand mai multi (Tabel 5.9). Totusi, prezenta intronilor nu este obligatorie existand si gene lipsite de acestia. Genele care codifica pentru moleculele de ARN functionale cum sunt ARNt si ARNr pot sa posede si ele secvente intercalare, dar intreruperile la nivelul acestora sunt mai putin frecvente. In general, cantitatea de ADN prezenta in introni depaseste cu mult pe cea din exoni.
ii) anumite gene apar de mai multe ori intr-un genom eucariot si contribuie la cresterea taliei. Existenta de copii multiple ale unei anumite gene nu este insa o proprietate exclusiva a eucariotelor. De exemplu, E. coli poseda sapte gene pentru ARNr, probabil pentru a satisface necesitatile, in ribozomi, ale unei cresteri rapide. Eucariotele poseda si ele gene in copiii multiple pentru a rezolva astfel de probleme.
La mamifere exista mai multe sute de gene care codifica pentru ARNr. O alta parte importanta din excesul de ADN este determinat de prezenta copiilor nefunctionale ale genelor. In general, aceste copii sunt numite pseudogene si sunt inactive din cauza existentei unor deletii sau altor alterari ale secventei ADN. Numarul pseudogenelor pentru un anumit tip de gena variaza foarte mult. Astfel, in genoamele eucariotelor, o gena poate fi prezenta o singura data, sau foarte bine, poate sa faca parte dintr-o familie de gene functionale strans inrudite. Acestea pot fi exprimate in momente diferite sau in tesuturi si celule diferite. Astfel de familii pot include si pseudogene. In figura 5.24 este prezentat clusterul b-globinei care contine doua pseudogene. De asemenea, se remarca cantitatea mare de "ADN nefunctional" prezenta la om comparativ cu drojdia Saccharomyces cerevisiae.
iii) o alta cauza a marimii excesive a genoamelor eucariote este prezenta unor familii mari de secvente de ADN repetitiv a caror functie este, pentru moment, necunoscuta. Astfel de familii reprezinta adesea intre 10% la 50% din genom (Tabel 5.10) . Aceste secvente au fost pentru prima data identificate in urma analizei cineticii de reactiei de renaturare a ADN. Astfel, s-a demonstrat ca mari cantitati de ADN se reasociaza mult mai rapid decat era prevazut pentru secventele unice. Viteza mare de reasociere indica faptul ca genoamele contin segmente care se repeta de sute de mii sau de milioane de ori (Figura 5.25) Curbele de reasociere Cot sunt informative pentru identificarea regiunilor care contin secvente repetitive sau sateliti. Clonarea moleculara si secventializarea au confirmat existenta unor astfel de secvente inalt repetitive, care, asemenea genelor repetitive, sunt aranjate in tandem sau sunt dispersate la nivelul a numeroase locusuri necorelate.
ADN repetitiv consta in secvente scurte repetitive sub forma unor copii identice sau inrudite in genom. Componenta care se renatureaza cel mai rapid in genomul eucariotelor este reprezentata de secventele "inalt repetitive", si consta in secvente foarte scurte care se repeta de multe ori in tandem formand "clustere" mari. Datorita unitatilor repetitive scurte, aceste secvente de ADN sunt adesea denumite secvente ADN simple" sau "secvente satelit"
Tabel 5. Variabilitatea numarului de introni si exoni si marimea acestorala diferite specii
|
Produsul genei |
Organism |
Exoni |
Introni |
|
|
Pb total |
numar |
Pb total |
||
|
Adenozin dezaminaza |
om | |||
|
Apolipoproteina B |
om | |||
|
Beta-globina |
soarece | |||
|
Citocrom b |
drojdii | |||
|
Dihidrofolat reductaza |
soarece | |||
|
Eritropoetina |
om | |||
|
Factorul VIII |
om | |||
|
Hipoxantin fosforibozil transferaza |
soarece | |||
|
Interferon alfa |
om | |||
|
Receptor LDL |
om | |||
|
Fazeolina |
Fasole verde | |||
|
Tiroglobulina |
om |
>40 | ||
|
ARNtTyr |
drojdii | |||
|
Ureaza (subunitate) |
soia | |||
|
Vitelogenina |
broasca | |||
|
Zeina |
porumb |
Tabel 5. . Tipuri de secvente ADN repetitive si distributia acestora
|
Secvente repetitive in tandem (unitatile repetitive la nivelul unui situs sunt de obicei identice) Proportia in genomul mamiferelor 10-15% Lungimea fiecarei unitati repetitive 1-10 pb Lungimea totala a repetitiilor ADN pentru un situs in perechi de baze: Sateliti ADN normali 100 000-10 milioane Minisateliti 100-100 000 Microsateliti 10-100 |
|
Secvente repetitive dispersate (copiile sunt similare dar nu identice) Proportia in genomul mamiferelor 25-40% Lungimea fiecarei unitati repetitive 100-10 000 pb Numar de repetitii in genom 1 la 1 milion |
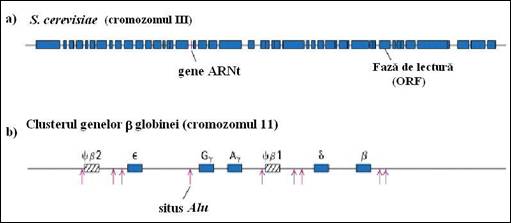
Figura 5.24. Prezentare comparativa a structurii genice a cromozomului III de la Saccharomyces cerevisiae (a) si clusterului b globinei de la om situat pe cromozomul 11(b). Dreptunghiurile albastre marcheaza fazele de lectura. Se poate constata ca genomul de S. cerevisiae are un numar foarte mic de secvente nefunctionale. La om clusterul globinei contine secvente intergenice necodante, pseudogene (yb1 si yb2) si secvente repetitive dispersate lungi (secventele Alu care apartin tipului SINES) (dupa Molecular Cell Biology Lodish H., et. al., 2000)
Figura 5. 25. Curbele de reasociere (Cot) pentru ADN dublu catenar din sursele indicate pe grafic. Se observa procesul de renaturare rapida a secventelor repetitive sau care contin sateliti dupa Molecular Cell Biology Lodish H., et. al., 2000)
Acest tip de secvente sunt prezente in aproape toate genoamele eucariote, dar cantitatea lor totala este extrem de variabila. In genoamele mamiferelor acestea reprezinta, in mod normal, < 10% dar la Drosophila virilis, cantitatea este de
Secventele repetitive in tandem pot fi corelate cu modificarea alinierilor care apar in urma imperecherii cromozomilor. Astfel, marimea "clusterelor in tandem" tinde sa fie foarte polimorfa, cu variatii largi intre indivizi. De fapt, clusterele mai mici, formate din astfel de secvente, pot fi folosite pentru caracterizarea genoamelor individuale folosind tehnica "fingerprinting" sau "genotiparea automata". Compararea secventelor simple de ADN, de la diferite specii, ofera informatii interesante despre mecanismele procesului de evolutie. Pentru moment nu putem spune cu precizie daca aceste secvente au o functie structurala.
Mari familii de repetitii dispersate in genomul mamiferelor
Analiza clasica a profilelor de dispersie demonstreaza ca genomul uman si genoamele altor mamifere poseda profile de dispersie apropiate. Aceste concluzii sunt confirmate de detaliile moleculare furnizate prin analiza de restrictie, clonare si secventializare. Cu toate ca exista diferite familii si sub-familii de repetitii dispersate, un numar mic dintre acestea prezinta un numar foarte mare de copii si domina genoamele. O familie dominanta poate reprezenta mai mult de 5% din ADN total, iar totalitatea familiilor pot reprezenta 20% din acesta. Unele familii sunt constituite din secvente de 100 la 500 pb si sunt denumite "secvente SINES Short Interspersed Repeats). Altele, ale caror membri pot avea 6 kpb sau mai mult, sunt denumite LINES Long Interspersed Repeats). Organisme cum sunt miomicetele, lacustele, echinodermele, pestii si amfibienii contin repetitii dispersate care se aseamana cu SINES de la mamifere. La plante si la nevertebrate exista secvente comparabile cu LINES de la mamifere.
Fiecare specie (ordin) poseda "un joc" caracteristic de familii de secvente SINES. Una dintre familiile SINES cele mai bine studiate este familia ALU de la primatele lumii vechi, denumita asa datorita prezentei in aceasta secventa a unui situs de restrictie Alu I. Unitatile Alu se gasesc in regiuni care flancheaza genele, in introni, in ADN satelit si grupate cu alte secvente repetitive dispersate (Figura 5.24
Pe langa multiplele familii SINES, genoamele mamiferelor poseda toate o abundenta familie LINES, familia LINE-1, ale carei membri pot avea 6 la 7 kpb. In unele dintre aceste genoame, exista si alte familii LINE dar ele nu sunt la fel de frecvente. Familiile LINE-1 de la diferite mamifere sunt toate inrudite dar divergente: cu cat speciile sunt mai apropiate cu atat familiile sunt mai similare. Secventele LINE-1 sunt prezente in regiunile care flancheaza genele, in introni si inserate in repetitiile ADN satelit.
Genoamele organitelor din celulele eucariote
Majoritatea celulelor eucariote poseda genoame mitocondriale si toate eucariotele capabile de fotosinteza au genoame in structura cloroplastelor. Initial, s-a considerat ca genoamele organitelor sunt molecule de ADN circulare. Aceasta deoarece hartile genetice si fizice obtinute, prin analiza de restrictie a genoamelor din mitocondrii si din cloroplaste, erau circulare. In anumite organite, studiile de microscopie electronica au evidentiat atat molecule circulare cat si liniare. La inceput, s-a presupus ca formele liniare ar putea reprezenta fragmente ale genoamelor circulare care s-au rupt in procesul de preparare. Din 1998 a fost admis ca unele eucariote inferioare (Paramecium, Chlamydomonas si multe suse de drojdii) poseda structuri lineare la nivelul genoamelor mitocondriale.
Surprinzator, marimea moleculelor de ADN mitocondrial (ADNmt), numarul si natura proteinelor pe care acestea le codifica, si chiar codul genetic mitocondrial, variaza mult intre diferitele tipuri de organisme (Tabel 5.11 si 5.12
Majoritatea animalelor multicelulare au genoame mitocondriale mici cu o organizare genetica compacta, genele fiind separare numai de spatii foarte reduse. ADNmt uman este tipic pentru aceasta categorie. Genomul mitocondrial este o molecula circulara care a fost complet secventializata si care contine 16 569 perechi de baze, fiind printre cele mai mici molecule de acest tip. Acesta codifica pentru doua molecule de ARNr prezente in ribozomii mitocondriali si pentru cei 22 ARNt folositi in procesul de translatie de la acest nivel. ADNmt uman contine 13 secvente care incep cu codonul ATG (metionina) si se termina cu un codon STOP. Aceste faze de lectura, destul de lungi pentru a codifica polipeptide de aproximativ 50 aminoacizi, au fost identificate cu proteinele codificate. ADNmt de la mamifere, spre deosebire ADN nuclear, este lipsit de introni si nu contine secvente necodante (Figura 5.26
ADNmt de la nevertebrate are aproximativ aceeasi marime cu cel uman, in timp ce ADNmt de la drojdii este de aproximativ de 5 ori mai mare (78 000 pb). Moleculele de ADNmt de la drojdii si de la alte eucariote superioare poseda gene care la mamifere sunt dispuse in nucleii celulelor (Figura 5.26
In contrast cu alte eucariote, care contin un singur tip de ADNmt, plantele contin mai multe tipuri de ADNmt care par sa se recombine unele cu altele. Aceste molecule de la plante sunt mult mai mari decat la alte organisme. Se poate intampla ca la o singura specie sa existe diferente foarte mari in marimea ADNmt (la pepenele verde 330 000 pb; la pepenele galben 2 500 000 pb).
Tabel 5. Marimea genoamelor mitocondriilor si cloroplastelor
|
Specie |
Tip de organism |
Marime genom (kb) |
|
A. Genoame mitocondriale Chlamydomonas reinhardtii Mus musculus Homo sapiens Metridium senile Drosophila melanogaster Chondrus crispus Aspergillus nidulans Reclinomonas americana Saccharomyces cerevisiae Suillus grisellus Brassica oleracea Arabidopsis thaliana Zea mays Cucumis melo |
Alga verde Vertebrate (soarece) Vertebrate (om) Nevertabrate Nevertebrate (musca) Alga rosie Ascomicete, fungi Protozoar Drojdie Bazidiomicete, fungi Plante cu flori (varza) Plante cu flori Plante cu flori (porumb) Plante cu flori (pepene) | |
|
B. Genoame ale cloroplastelor Pissum sativum Marchantia polymorpha Oryza sativa Nicotiana tabacum Chlamydomonas reinhardtii |
Pante cu flori Plante Plante cu flori (orez) Plante cu flori (tutun) Alga verde |
Figura 5. Genoamele mitocondriale de la om (a) si de la Saccharomyces cerevisiae
Spre deosebire de animale, drojdii si fungii, ADNmt de la plante contine gene care codifica pentru ARNr 5S, care este prezent numai in ribozomii mitocondriilor plantelor, si pentru subunitatea a a ATP-azei F1. ARNr de la plante este mult mai mare decat cel de la alte eucariote. Secventializarea recenta a uneia dintre cele mai mici molecule de ADNmt de la plante a evidentiat ca marimea este datorata existentei de regiuni necodante si de secvente duplicate.
Diferentele de marime si de capacitate codanta a ADNmt de la diferite organisme poate reflecta un anumit schimb de ADN, intre mitocondrie si nucleu, in cursul evolutiei. Dovezi directe, in favoarea acestei afirmatii au fost furnizate de evidentierea unor proteine ale caror gene sunt la unele specii localizate in nucleu si la altele in mitocondrie. Se pare ca este vorba de o deplasare a genelor din mitocondrie in nucleu si invers, in timpul evolutiei. Cel mai elocvent exemplu este cel al genei cox II, care codifica subunitatea 2 a citocrom c oxidazei. Pentru toate organismele studiate, aceasta gena se gaseste in ADNmt, cu exceptia unei specii de legume numita Bob (mung bean) unde gena cox II este nucleara.
Tabel 5. Caracteristicile genelor mitocondriale
|
Caracteristica |
C. reinhadtii |
Homo sapiens |
S. cerevisiae |
A. thaliana |
R. americana |
|
Numar total de gene Tipuri de gene Gene care codifica proteine complexul respirator proteine ribozomale proteine transportoare ARN polimeraza Factor de translatie Gene care codifica ARNr Gene ARNt Alte gene ARN Numar de introni Marimea genomului (kb) |
Multe molecule de ARN transcrise de la plante sunt procesate prin actiunea unei enzime care catalizeaza transformarea unor resturi selectionate C in U si ocazional a U in C. Gena nucleara cox II de la bob este mai apropiata structural de moleculele de ARN procesate decat de gena mitocondriala cox II prezenta la alte legume. Aceste date sunt dovezi ale deplasarii genei cox II din mitocondrie in nucleu printr-un proces care implica participarea unei molecule de ARN. Este posibil un mecanism de reverstranscriptie similar celui prin care pseudogenele procesate sunt generate in genomul nuclear prin intermediul ARNm.
Structura cloroplastelor este similara in multe privinte cu aceea a mitocondriilor. Ca si acestea, cloroplastele contin mai multe copii de ADN si ribozomi care codifica pentru proteine din cloroplaste, folosind codul genetic standard. Alte proteine specifice din cloroplastelor sunt fabricate in citoplasma si sunt transportate, dupa traducere.
ADN din cloroplaste sunt molecule circulare de aproximativ 120 000-160 000 pb, in functie de specie. Multe dintre moleculele de ADN din cloroplaste au fost complet secventializate (Tabel 5.11). Genomul cloroplastelor din planta a carei denumire populara este "plaman" prezinta doua secvente repetitive inverse fiecare constand in 10058 pb care contin genele ARNr si alte cateva gene duplicate. In ciuda diferentei de marime (plaman - 121 024 pb; tutun - 155 844), intreaga organizare si compozitia genelor de la plaman si de la tutun este foarte similara; Diferenta de marime apare, in primul rand, de la repetitiile inversate provenite din duplicarea anumitor gene.
Din aproximativ 120 de gene codificate de ADN din cloroplaste, aproape 60 sunt implicate in transcriptia si translatia ARN si includ genele care codifica pentru ARNr, ARNt, subunitatile ARN polimerazei si proteinele ribozomale. Aproximativ 20 de gene codifica pentru subunitatile complexelor transportoare de electroni din cloroplast si pentru complexul FoF1 ATP-azic. De asemenea, sunt codificate de catre genomul cloroplastelor cele doua subunitati mari ale ribulozo 1,5 difosfat carboxilaza, care este implicata in fixarea dioxidului de carbon in timpul fotosintezei.
Reflectand originea endosimbiotica a cloroplastelor unele regiuni din genomul acestora sunt similare cu cele ale ADN de la bacteriile actuale. De exemplu, ADN din cloroplaste codifica patru subunitati ale ARN polimerazei care sunt omologe cu subunitatile ARN polimerazei de la E. coli. Un alt segment al ADN din cloroplaste codifica 8 proteine care sunt omologe cu 8 proteine ribozomale de la E. coli. Ordinea genelor este similara in cele doua tipuri de ADN. ADN din cloroplastele plamanului poseda cateva gene care nu sunt in ADN din cloroplastele tutunului si invers. Acest lucru poate indica un posibil schimb ancestral intre cloroplaste si nucleu.
|
Politica de confidentialitate |
| Copyright ©
2024 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |