1771 - Anatomistul britanic William Hewson descrie primele cercetari referitoare la coagularea sangelui, reusind sa izoleze ceea ce el a denumit "limfa coagulanta", echivalentul fibrinogenului.
1901 - este anul in care Karl Landsteiner va publica descoperirea grupelor sanguine, initial denumite A,B si C, care ulterior va deveni O. Va defini in urma rezultatelor experimentale notiunea de reactie de aglutinare, anticorpi si aglutinie.
1902 - Karl Landsteiner alaturi de Alfred von Decastello si Adriano Sterly finalizeaza studiile si experimentele practice referitoare la caracteristicile grupei sanguine AB.
1914 - Doctorul Albert Hustin din Belgia va demonstra proprietatile anticoagulante ale citratului de sodiu.
1916 - Francis Peyton Rous si J.R. Turner vor demonstra ca o solutie de glucoza-citrat de sodiu poate asigura pastrarea sangelui dupa recoltare pentru o durata de cateva saptamani, descoperire ce va salva multe vieti in primul razboi mondial.
Dupa fundamentarea stiintifica a compozitiei sangelui au fost validate metode care vizeaza determinarea unor categorii de proteine fixate fie pe membrana eritrocitara sau existente in serul sangvin, care discrimineaza indivizii in diverse categorii. De aceste metode sunt legate numele unor medici, fiziologi, unele purtandu-le numele : metoda Beth-Vincent, Simonin, Holtzer etc.
Stiinta se va dezvolta rapid, iar descoperirile biologiei moleculare vor deschide o noua era, cea a geneticii.
- J Watson si F. Crick sunt cei doi savanti care prin difractia razelor X au stabilit structura de dublu helix a macromoleculei de ADN, motiv pentru care in anul 1962 li s-a decernat premiul Nobel.
1985 Hegemonia amprentelor papilare a fost egalata prin progresele recente inregistrate de biologia moleculara prin folosirea celui mai discriminativ criteriu biologic, molecula ADN (acidul deoxiribonucleic), purtatoarea codului genetic. Metoda a fost adaptata scopurilor judiciare de catre geneticianul Alex Jeffreys si are ca fundament stiintific hipervariabilitatea genomului uman, ce determina unicitatea individului.
Initial a fost utilizata metoda R.F.L.P. (restriction fragment lenght polymorfism), ce are ca principiu polimorfismul lungimii fragmentelor de restrictie prin scindarea macromoleculei de A.D.N. cu ajutorul enzimelor de restrictie, vizualizarea acestor fragmente si compararea profilelor obtinute.
Aceasta metoda este laborioasa, materialul genetic analizat trebuie sa fie integru si in cantitate relativ mare, durata determinarilor considerabila, vizualizarea utilizand markeri radioactivi .
1993 - Karry Mullis, laureat al premiului Nobel fundamenteaza metoda 'reactiei in lant a polimerazei' (Polymerase Chain Reaction), prin care este posibila multiplicarea de miliarde de ori a cantitatii de A.D.N. dintr-o urma. Procesul de amplificare permite investigarea materialului biologic partial degradat, in acord cu realitatea.
Individualizarea, aproape certa a urmelor biolgice prin genotipare,a determinat in mod gresit sa i se atribuie prin similitudine cu unicitatea impresiunii papilare, acestei metode denumirea de "amprenta genetica", termenul stiintific unanim acceptat fiind acela de profil genetic.
Prin urme biologice, intr-o acceptiune globala, se pot defini totalitatea urmelor alcatuite din organisme biologice (microorganisme, plante, animale) sau componente ale acestora (organe, tesuturi).
Într-o acceptiune restransa urmele biologice de interes criminalistic le-ar grupa numai pe cele umane (sange, secretii, diverse tipuri de tesuturi, fire de par sau celule).
Abordarea ar fi limitata excluzandu-se situatiile in care urmele de sol ce contin microorganisme cu areal foarte restrans (soluri saraturoase sau acvatic), larvele insectelor, fragmentele vegetale, polenul, firele de par animal, pot constitui dovezi care pot proba prezenta suspectilor la locul infractiunii, sau participarea acestora la acte de braconaj, sau uneori timpul scurs de la deces.
Microorganismele si fungii
Sunt cei mai importanti factori distructivi ai urmelor biologice, prin actiunea fermentativa anaeroba si atacul propriilor enzime proteolitice vor metaboliza mediile proteice si polizaharidice. Prin aceasta degradare sunt distruse proteinele, glucidele si lipidele celulare, actiunea indelungata distrugand in totalitate chiar acizii nucleici.
În general, fiind organisme anaerobe, un factor favorizant il constituie ambalarea urmelor biologice in recipienti ermetic inchisi.
Umezeala, caldura si radiatiile ultraviolete
Sunt conditii optime pentru dezvoltarea microorganismelor si fungilor, motiv pentru care este necesara uscarea imediata a urmelor biologice si ambalarea acestora in hartie.
Caldura excesiva poate duce la denaturarea ireversibila a proteinelor si acizilor nucleici, orice determinare fiind compromisa.
Radiatiile ultraviolete (radiatiile solare) pot fi un factor distructiv al fragmentelor de acizi nucleici. Expunerea probelor biologice la radiatiile UV pentru un timp indelungat poate modifica ireversibil structura A.D.N.
Actiunea substantelor chimice
Acizii, bazele, colorantii, substantele puternic oxidante sau reducatoare actioneaza distructiv prin denaturarea ireversibila a structurii proteinelor.
Enzimele celulare
Sunt localizate in citoplasma sau nucleul celular fiind responsabile de actiunea proteolitica ce duce la scindarea proteinelor citoplasmatice in fragmente peptidice, in final obtinandu-se un mediu de cultura optim atacului bacterian.
O clasa aparte de enzime o reprezinta endonucleazele de restrictie a caror actiune scindeaza lantul dublu catenar al acidului deoxiribonucleic cu formarea de fragmente degradate ce fac imposibila genotiparea urmei biologice.
Actiunea proteolitica a enzimelor poate fi limitata prin uscarea urmelor biologice, urmata de pastrarea la temperaturi cuprinse intre +4 si - 10oC.
Probele biologice sunt utilizate pentru probarea contactului fizic al unei persoane cu o alta persoana, cu un obiect, sau prezenta unei persoane intr-o anumita locatie. Fenomenul prin care urmele biologice sunt preluate pe diverse suporturi se numeste fenomen de transfer.
Urmele biologice umane (sange, sperma, fire de par etc.) pot fi transferate in mod direct pe corpul unei persoane, pe imbracamintea acesteia, sau pe un obiect gasit la fata locului. Depuse pe suporturi, lichidele biologice adera sub forma de pete. Urmele biologice solide (tesuturi, fire de par) se pot depune pe obiecte prin contact direct.
Transferul urmelor biologice poate fi rezultatul urmatoarelor situatii:
a. Transferul direct (primar)
o Transferul produsilor biologici ce apartin suspectului pe victima (pe corpul acesteia sau pe obiectele sale de imbracaminte).
o Transferul produsilor biologici ce apartin suspectului pe un obiect din campul infractional.
o Transferul produsilor biologici ce apartin victimei pe suspect (pe corpul acestuia sau pe obiectele sale de imbracaminte).
o Transferul produsilor biologici ce apartin victimei pe un obiect din campul infractional.
o Transferul produsilor biologici ce apartin unui martor pe victima sau suspect.
o Transferul produsilor biologici ce apartin unui martor pe un obiect din campul infractional.
b. Transferul indirect (secundar)
Sunt cazuri in care produsii biologici pot fi transferati victimei, suspectului sau pe un obiect, printr-un vector intermediar. În acest caz transferul este secundar si nu exista contact fizic nemijlocit intre sursa si suport. Vectorul poate fi o persoana sau obiect. Transferul indirect al urmei nu constituie proba pentru a lega o anumita persoana de infractiune.
Avand in vedere perisabilitatea urmelor biologice sub actiunea factorilor distructivi, recoltarea si ambalarea urmelor ridicate din campul infractional devin hotaratoare in probatoriul unei cauze. Daca probele nu sunt recoltate, inregistrate, ambalate si conservate corespunzator, nu vor fi admise in justitie, sau degradarea biologica nu va permite obtinerea unor rezultate valorificabile. Etapele ce se succed de la identificare si recoltare pana la receptia acestora in laborator poarta denumirea "de lant al posesiunii".
Cercetarea campului infractiunii se va efectua obligatoriu folosind manusi, care se vor schimba ori de cate ori se trece la atingerea si ridicarea unei noi probe.
Pentru manipularea probelor se va folosi instrumentar steril. Nu se vor manipula simultan mai multe probe.
Probele nu vor veni in contact una cu alta, ambalandu-se separat. Se va realiza o separare intre probele provenite de la suspect si cele de la victima, in vederea evitarii intercontaminarii.
Nu se vor atinge cu mana descoperita obiectele in litigiu, se va evita vorbitul, stranutatul sau tusitul spre obiectele purtatoare de urme.
Pe timpul recoltarii probelor se va evita fumatul sau consumul de alimente.
Pe cat posibil se vor trimite laboratoarelor specializate obiectele purtatoare ca atare, fara a incerca prelevarea sau decuparea urmelor identificate sau presupuse a fi urme biologice.
Probele se vor usca prin expunere la aer inaintea ambalarii, iar ambalarea se va efectua in ambalaje de hartie noi, fiind interzisa ambalarea in pungi din material plastic sau sticla inchise ermetic (evitarea fenomenului de condensare, umidificare, atac bacterian anaerob)
Nu se vor ambala mai multe obiecte in acelasi container.
Serologia judiciara a constituit o etapa "clasica" in istoria criminalisticii ce a raspuns unor exigente ale momentului.
Polimorfismul structural al proteinelor reprezinta o caracteristica generala a lumii vii, proprietate pe care se fundamenteaza multe din metodele criminalistice.
Configuratiile structurale sunt comune diferitelor specii de animale, astfel incat unele grupe sau sisteme complete de grupe sunt prezente la anumite specii animale, mai ales la mamiferele superioare.
În general notiunea de grup sanguin, are in vedere sistemul AB0 descoperit in 1901 de K. Landsteiner. Antigenele din sistemul de grupe AB0 sunt insa cele mai raspandite in regnul animal, fiind prezente chiar la unele microorganisme (grupa A).
Notiunea de "grupa sanguina" cuprinde totalitatea sistemelor grupale rezultate din polimorfismele structurale biochimice, indiferent daca ele sunt localizate pe membrana celulelor sanguine, in proteinele plasmatice sau pe celulele ori limfa celulara din diferitele tesuturi si organe.
În cazul omului, antigenele sunt prezente intr-o proportie de 80 in toate organele, in unele secretii, conferind persoanei "caracter secretor", restul proportiei de 20 avand statut de persoane "nesecretoare". Alaturi de factorii de grupe AB0, in fiecare caz este prezenta si antigena H, care diferentiaza sangele uman de cel animal, aceasta existand si la persoane cu grupa 0. Chiar Landsteiner si cercetarile ulterioare au demonstrat faptul ca pe membrana eritrocitului si a altor celule hematice sunt numeroase variatii individuale cu caracter antigenic, constituente ale sistemelor de grupe independente.
Cercetarile, prin metoda gel-electroforezei, au demonstrat prezenta in serul sanguin a izoenzimelor specifice de grup, denumite "grupe serice" care sunt expresia altor tipuri de variabilitate.
Apartenenta indivizilor la anumite grupe sanguine, grupe de izoenzime serice GPM (fosfoglucomutaza), EAP (Fosfataza acida eritrocitara), GC (Componentul de grup specific) isi gaseste aplicabilitatea in serologia judiciara, prin determinari comparative ale urmelor biologice ridicate din campul infractional cu standarde ale caror caracteristici sunt foarte bine cunoscute, in final atribuindu-se in cazul unor reactii identice, proprietatile comune celor doua tipuri de probe.
Aceasta discriminare este relativa si limitata, statisticile demonstrand ca in general 70-80 din indivizi apartin grupelor sanguine 0I si AII, 10-12 grupei sanguine BIII, iar restul grupei sanguine ABIV, deci este evident ca situatia in care victima si banuitul pot apartine aceleiasi grupe sanguine este foarte frecventa.
A. Urmele de sange
Identificarea urmelor de sange depuse pe suporturi, sau in stare lichida se realizeaza prin reactii de orientare si reactii de certitudine.
Reactiile de orientare se bazeaza pe proprietatile componentelor sangelui care prin modificare chimica in prezenta unor cromofori duc la aparitia unor culori specifice. Reactiile de orientare au un mare avantaj fiind foarte rapide si sensibile, dar prezinta un dezavantaj din punct de vedere al specificitatii, existand posibilitatea unor interferente. Lumea stiintifica a acceptat principiul prin care daca mai multe reactii de orientare, sunt pozitive, de obicei combinatii intre cele deosebit de sensibile si cele foarte specifice, se considera demonstrata natura acestor urme. Dintre cele mai utilizate reactii de orientare de laboratoarele criminalistice se pot enumera: reactivul Adler, reactivul Guarino, reactivul Kastl-Mayer etc.
Reactiile de certitudine au ca principiu demonstrarea prezentei hemoglobinei, sunt mai costisitoare, necesitand un timp mai indelungat pentru determinari. Cele mai utilizate sunt reactia Taichmann, reactia Gabrielli - Bertrande sau reactia Takayama. Aceste metode sunt completate de examinarea microspectroscopica, caz in care urmele de hem prezinta 2 benzi de absorbtie in zona lungimilor de unda de 589 -577 nm si 556 - 536 nm.
Reactiile de orientare si probabilitate
Se bazeaza pe proprietatile biochimice, enzimatice si imunologice ale lichidului spermatic.
Pentru suporturile cu suprafata mare una din cele mai uzitate metode o reprezinta examinarea suprafetelor mari prin examinare in lumina U.V., in care urmele de sperma prezinta fluorescenta, specifica insa si altor substante.
Dintre reactiile de probabilitate, cele mai des folosite sunt cele bazate pe reactiile cristalografice, enzimatice si antigenice ale lichidului seminal.
Reactiile cristalografice au ca principiu obtinerea de cristale specifice ca urmare a reactiei dintre componentele anorganice si cele ale lichidului seminal. (r. Florance, Barberino, Bocarius etc.) Limitele acestor determinari sunt generate de cantitatea mare de proba necesara.
În practica cele mai utilizate metode sunt cele ce se bazeaza pe activitatea fosfatazei acide a lichidului seminal (r. Phosphatesmo) si reactia de determinare a antigenelor specifice din prostata (glicoproteine intracelulare cu masa moleculara de 3400 Dalton, sintetizate numai de catre glandele prostatice - r. P.S.A. Check).
Reactiile de certitudine
Indiferent de rezultatul testelor de orientare, se efectueaza frotiuri pentru observarea directa a prezentei sau absentei spermatozoizilor. Pentru vizualizare se folosesc diverse sisteme de colorare (hematoxilin-eozina, eritrozina etc.) care au drept scop colorarea diferentiala a materialului nuclear de cel citoplasmatic, fiind foarte bine definite criteriile de identificare (forma, marime, coloratie).
C. Urmele de saliva
Sunt in general greu detectabile la o examinare preliminara, in lumina ultravioleta emitand o fluorescenta slaba.
Reactia de identificare se bazeaza pe vizualizarea actiunii amilazei, care in prezenta unui substrat de amidon inalt polimerizat colorat in albastru va elibera in reactie colorantul.
D. Urmele de natura piloasa
Se recolteaza impreuna cu alte fibre depuse pe suporturi, atribuirea naturii(fibre vegetale, textile, animale sau umane) se realizeaza numai prin examinarea microscopica intre lama si lamela. Examinarea microscopica urmareste observarea caracteristicilor morfologice (canal medular, cuticula, secventa de culoare, grosime si bulb) Aceste examinari aduc informatii in vederea stabilirii speciei, iar in cazul firelor de par umane se poate indica zona de provenienta, modul de detasare, elementele de asemanare sau diferentiere intre firele de par litigiu si cele model de comparatie.
Pentru stabilirea speciei urmelor biologice se utilizeaza un control pozitiv constituit din sange uman, sau al diverselor specii de animale, in cazul reactiilor identice ale litigiului si ale controlului pozitiv demonstrandu-se ca ambele probe provin de la aceeasi specie.
Reactia de precipitare (Uhlenhut - Cistoreici - Wassermann).
La limita de jonctiune dintre maceratul urmei de sange si serul precipitat se formeaza o pelicula fina (inel) de precipitat albicios.
În cazul urmelor de sperma, pentru determinarea speciei se utilizeaza serul anti-sperma umana.
Reactia hartmann toilliez (de difuziune in gel).
Reactia de imunoprecipitare este realizata prin difuziunea spontana a proteinelor (antigene) fata de anticorpii din serul precipitant. Proteinele din urme se dizolva si difuzeaza in geloza in mod circular. Totodata difuzeaza si anticorpii din serurile de testare, iar la frontul de intalnire al antigenelor cu anticorpii specifici va produce o linie (sau 2-3 linii) de precipitare. Aceste linii sunt caracteristice.
Testul imunocromatografic (Hexagon OBTI)
se bazeaza pe detectia imunologica a hemoglobinei, sensibilitatea metodei fiind de 0,05µg/ml.
Posibilitatea de identificare a grupelor sanguine din urmele biologice se bazeaza pe principiul activitatii biochimice a antigenelor prezente pe membrana eritrocitelor si a anticorpii naturali serici, atat in cazul urmelor biologice lichide cat si in cazul urmelor biologice depuse pe suporturi, activitatea lor fiind dependenta de expunerea la factorii de mediu.
Metoda pentru identificarea grupelor sanguine din urmele biologice va fi adaptata intotdeauna la conditiile concrete: vechimea, gradul de uscare, gradul de alterare si suportul urmelor (daca acesta este absorbant sau nu, daca are influenta asupra reactiei antigen-anticorp).
Pe membrana eritrocitelor se constata prezenta sau absenta aglutinogenelor A si B, individual sau impreuna. În serul sanguin se identifica aglutininele α si β.
Metodele de determinare a grupei sanguine pun in evidenta prezenta, sau absenta aglutinogenelor si aglutininelor.
În cazul sangelui lichid, stabilirea grupei se efectueaza pe baza reactiei dintre serurilor de testare din grupele anti - A,B,O cu suspensia de eritrocite neidentificate (metoda Beth - Vincent), respectiv prin reactia serului litigiu cu suspensie de eritrocite A si B (metoda Simonin). in criminalistica se utilizeaza ambele procedee simultan.
- Identificarea aglutininelor
Se poate realiza prin metoda Lattes, atat pentru urmele de sange cat si pentru secretii (sperma, secretie vaginala, saliva)
- Identificarea antigenelor
Se realizeaza prin metoda absorbtiei-elutiei, metoda absorbtiei si prin metoda Holtzer.
Examinarea comparativa reprezinta un principiu stiintific prin care o proba litigiu este examinata simultan cu o proba de referinta (model de comparatie), in vederea determinarii elementelor comune.
Demonstratia este etapa prin care totalitatea informatiilor acumulate in etapele examinarilor fixeaza caracteristicile probelor analizate. Pentru demonstrarea concluziilor pot fi utilizate fotografii, sau orice alta documentatie aferenta.
Concluzia pozitiva
Reprezinta certitudinea determinarilor efectuate, al caror rezultat a fost pozitiv (identic cu cel al controlului), pentru stabilirea naturii urmei biologice (sange, saliva, fire de par, lichid seminal, secretie vaginala etc.), pentru determinarea provenientei umane sau animale a urmelor si pentru determinarea apartenentei acestora la un anumit grup sanguin.
Concluzia negativa
Reprezinta certitudinea analizelor al caror rezultat a fost negativ, determinand excluderea unor urme care desi au aspect de urme biologice, nu poseda aceasta calitate.
Concluzia de probabilitate
Reprezinta ilustrarea discordantelor partiale cu seria controalelor pozitive si negative ale reactiilor de determinare.
Exemple:
urma biologica determina reactie pozitiva numai in contact cu reactivul Adler, contactul cu reactivul Guarino neproducand reactie specifica (desi reactia controlului pozitiv este evidenta) - "urma este probabil de sange".
in contact cu serul anti-om maceratul urmei biologice produce un inel foarte subtire, greu vizibil, (desi reactia controlului pozitiv este evidenta) - "urma de sange este probabil umana".
Concluzia de imposibilitate
Se formuleaza in cazul lipsei generale de reactivitate datorate cantitatii foarte reduse de proba, al vechimii, al actiunii prelungite al factorilor de mediu, al impurificarii urmei cu produsi chimici etc.
Determinarea naturii, speciei si apartenenta urmelor biologice la un anumit grup sanguin. Pentru urmele de saliva si lichid seminal determinarea caracterului secretor/nesecretor.
Determinarea naturii, speciei si a caracteristicelor morfologice a firelor de par. Compararea acestor caracteristici pentru probele litigiu si model de comparatie in vederea stabilirii elementelor de asemanare si deosebire.
Performantele acestor metode constau in timpul relativ scurt de efectuare si costuri scazute. Nu necesita aparatura de laborator deosebita.
Limitele majore sunt datorate de puterea redusa de discriminare a urmelor, iar in cazul amestecurilor urmelor biologice nu se pot stabili grupele sanguine sau serice ale participantilor, ci numai cea a amestecului in totalitate.
Un alt dezavantaj il constituie cantitatile mari de probe biologice necesare determinarilor si caracterul de probabilitate al unor determinari datorat limitarii cantitative.
Proteinele, clasa de substante majoritara in determinarile serologice, sunt substante foarte sensibile fata de actiunea factorilor de mediu motiv pentru care posibilitatea compromiterii rezultatelor obtinute este foarte frecventa.
Informatia genetica a fiecarui individ este stocata in genom. Acesta este definit ca un set complet de cromozomi mostenit ca o unitate de la ambii parinti. În structura cromozomului sunt prezente genele care controleaza caracteristicile ereditare ale organismului. Pozitia ocupata de o gena in cromozom poarta denumirea de locus. Genele sunt molecule complexe constituite din acizi nucleici - ADN sau ARN in cazul unor virusi. Genomul uman este constituit din 22 de perechi de cromozomi autosomali si doi care determina sexul, astfel nucleul unei celule normale contine 46 de cromozomi diferiti sau 23 perechi, una care provine de la mama si una care provine de la tata.
Materialul genetic, ADN-ul din cromozomi este compus din secvente codante denumite exoni (portiuni care codifica sinteza proteinelor) si portiuni noncodante cunoscute sub numele de introni. Regiunile codante sunt cunoscute ca gene si contin informatia necesara pentru ca o celula sa fabrice proteinele. În genomul uman exista aproximativ 50000 - 100000 de gene.
Variabilitatea genetica a indivizilor este data de genele sau markerii regasiti la nivelul regiunilor noncodante ale genomului uman.
O fractiune mica din ADN-ul total (cca. un milion de nucleotide - unitati de baza ale acizilor dezoxiribonucleici) difera de la individ la individ, ceea ce face ca fiecare individ sa fie unic. Aceste regiuni variabile permit folosirea informatiei ADN pentru identificarea umana.
Au fost dezvoltate metode de localizare si caracterizare a variabilitatii genetice din aceste regiuni ale genomului uman.
Altfel spus, intronii sunt formati din blocuri repetitive de cca. 3-7 nucleotide care se repeta de un numar de ori, specific fiecarui individ.
În determinarea genotipului (profilului genetic) se folosesc mai multi markeri sau loci. Fiecare locus are un continut de alele, care poate fi definit ca unitati repetitive. Cu cat sunt determinati mai multi markeri cu atat mai mare este probabilitatea ca doi indivizi genetic diferiti sa posede genotipuri diferite. Invers, fiecare locus determinat suplimentar face sa creasca gradul de discriminare atunci cand se afirma ca doua probe avand aceleasi profile ADN provin de la acelasi individ.
Pe langa bazele geneticii moleculare, in determinarea si exprimarea profilului genetic al unei persoane sunt utilizate si anumite calcule biostatistice. Daca fiecare locus este mostenit independent de ceilalti loci atunci pentru a calcula frecventa unui anumit profil ADN se inmultesc frecventele tuturor genotipurilor care intra in profilul respectiv. Acest calcul este cunoscut sub numele de regula produsului.
Variabilitatea genetica a secventei ADN de pe tot globul este stocata intr-o baza de date computerizata, GenBank. GenBank este realizata de National Center for Biotechnology Information (NCBI), departament al National Library of Medicine din cadrul US Institutes of Health. NCBI a fost infiintat in anul 1988 ca institutie nationala pentru informare in domeniul biologiei moleculare in scopul imbunatatirii intelegerii proceselor moleculare legate de starea de sanatate si boli. Pana in august 1999 GenBank a acumulat 4,6 milioane de inregistrari secvente ADN, cuprinzand peste 3,4 bilioane de nucleotide. Peste 5000 de specii sunt prezente in GenBank. Secventele ADN din GenBank pot fi accesate din pagina NCBI, la adresa https://www.ncbi.nlm.nih.gov.
Din fiecare urma biologica umana identificata ca fiind de interes in rezolvarea sau aducerea unor informatii pentru rezolvarea unui caz, se extrage ADN-ul care este apoi purificat si amplificat. Metoda utilizata pentru amplificare este "Short Tanden Repet" (Fragmente Mici Repetitive) cuplata cu Poly Chain Reaction (Reactia de Amplificare in Lant) - care este un proces enzimatic prin care o anumita zona a ADN este replicata de mai multe ori cu formarea unui numar mare de copii ale unei anumite secvente ADN. Dupa 30 de cicluri se genereaza cca. un bilion (109) de copii ale zonei tinta din matrita ADN. Utilizandu-se reactivi specifici fluorescenti, produsii astfel amplificati sunt analizati prin electroforeza capilara.
Rezultatele obtinute sunt sub forma unor picuri grupate intr-o elelectroferograma, care ocupa anumite pozitii din cadrul unei scari ce contin toate alelele studiate. Fiecare astfel de pic poarta un anumit numar, obtinandu-se in final o combinatie de cifre.
Cea mai importanta caracteristica a acestei metode este obtinerea unor profile genetice corespunzatoare urmelor biologice din care a fost extras ADNul. În cazul a doua profile genetice identice recoltate din doua urme biologice diferite, se poate afirma ca au fost create de aceeasi persoana cu foarte mare probabilitate (aproape certitudine).
Dintre avantajele metodei pot fi notate:
Poate analiza ADN partial degradat.
Pot fi folosite cantitati foarte mici de matrita ADN provenind chiar si dintr-o singura celula - o cantitate minima de ADN de 0,2ng;
ADN-ul degradat pana la fragmente de cateva sute de perechi de baze poate servi drept matrita pentru amplificare.
Permite discriminarea amestecurilor de urme biologice ce provin de la persoane diferite.
Pot fi partial automatizate
Puterea de discriminare a indivizilor in cazul unor amestecuri de mai multe persoane este invers proportional cu cresterea numarului de profile aflate in amestec.
Probele biologice care au fost supuse timp indelungat actiunii apei, luminii solare, au fost degradate bacterian, sau cele in care probele biologice sunt in amestec cu inhibitori (coloranti, sol, materii grase etc.) pot limita analizele genetice, deoarece actiunea acestor factori pot inhiba procesul de multiplicare a fragmentelor de A.D.N. tinta.
Daca probele biologice ce urmeaza a fi analizate nu sunt recoltate conform unei strategii caracteristice se pot obtine rezultate neinterpretabile.
Din luna octombrie 2003, cand au fost incheiate cu succes testele de acceptanta care validau aparatura si tehnologia utilizata, functioneaza in cadrul Institutului de Criminalistica Laboratorul de genetica judiciara.
În paralel s-a desfasurat procesul de pregatire teoretica si practica a celor 8 specialisti, proces coordonat de dr. Frantz Neuhuber seful departamentului de genetica judiciara a Institutului Medico-Legal din Salzburg, alaturi de alti mari specialisti cum ar fi prof Harald Scheinder sau dr. Peter Wiegant.
Structural Laboratorul de genetica se compune din trei compartimente:
laborator de analiza urme ridicate de la fata locului;
laborator de analiza probe recoltate de la persoanele care au comis infractiuni cu violenta (pentru constituirea bazei de date) sau de la persoanele suspecte in cauza. Politia este prima institutie din tara care si-a propus sa constituie o baza de date genetice, fara de care nu poate fi conceputa interpretarea in domeniul judiciar a rezultatelor de tipare a ADN;
subsistem informatizat de prelucrare a rezultatelor din cele doua laboratoare si formare a bazei de date cu profilele ADN ale persoanelor care au comis infractiuni cu violenta.
Proiectul a avut ca prioritati asigurarea integrala a conditiilor tehnice pentru respectarea tuturor recomandarilor si normelor de calitate, privind amenajarea laboratoarelor, protocoalele de lucru, pregatirea personalului si practica de laborator (Good Laboratory Practice), conform standardelor cerute pentru biologia judiciara de organismele internationale (Technical Working Group on DNA Analysis Methods [TWGDAM], European DNA Profiling Group [EDNAP], International Society for Forensic Genetics [ISFG] ) fiind dotat inca din start cu logistica necesara (reactivi, consumabile, piese de schimb) pentru functionarea laboratoarelor timp de trei ani
În cadrul sistemului, fluxul de analiza a probelor ridicate de la fata locului este complet separat de fluxul de procesare a probelor de referinta. Se respecta astfel una dintre cerintele importante pentru prevenirea intercontaminarii probelor, pericol care exista atunci cand fluxurile de analiza se intersecteaza
Determinarea naturii urmelor biologice identificate si reactiile pentru stabilirea speciei sunt etape comune cu determinarile utilizate de serologia clasica.
Extractia va fi adaptata la tipurile de probe analizate:
izolarea ADN - ului din urmele biologice ridicate de la scena crimei-extractia organica cloroform/alcool izoamilic/fenol;
izolarea ADN - ului din probe de sange si probe bucale extractia cu rasina chelex;
izolarea ADN - ului din secretia vaginala extractia diferentiala cu ajutorul proteinazei K si DTT (ditiotreitol).
Cuantificarea ADN, esentiala pentru obtinerea unor rezultate finale valide, va urmari initial determinarea ADN - ului total, iar ulterior se va determina ADN - ul uman din probele analizate:
Cuantificarea ADN - ului total, fluorimetric, cu colorantul Hoechst 33258 (bisbenzimida): lexcit = 365 nm; lemisie
Cuantificarea ADN - ului uman (etape):
Ø amplificarea locusului FGA cu Termocycler PE 9700;
Ø electroforeza in gel de poliacrilamida;
colorare prin silver staining.
Amplificarea secventelor repetitive scurte (STR - Short Tandem Repeats) se va face numai in sisteme multiplex, cu primeri marcati fluorescent, sistemele folosite permitand o discriminare la nivel inalt a probelor, cu excluderea oricaror erori in stabilirea conditiilor de identificare a unei probe. Vor fi folosite doua sisteme multiplex:
AmplFISTR Identifiler Kit - 16 locusuri : CSF1PO, D3S1358, D5S818, D7S820, D8S1179, D13S317, D16S539, D18S51, D21S11, vWA, FGA, TH01, TPOX, D2S1338 and D19S433 si Amelogenin (patru markeri fluorescenti)
AmplFISTR SGM plus - 11 locusuri: D3S1358, D8S1179, D16S539, D21S11, vWA, D18S551, FGA, TH01, D2S1338 D19S433 si Amelogenin
Analiza produsilor PCR si obtinerea profilelor genetice ale probelor biologice se realizeaza automat, pe aparate moderne, de mare productivitate - secventiatoare livrate de firma Applied Biosystems - ABI Prism 310 (pentru probele ridicate de la fata locului) si ABI 3100 (pentru probele analizate pentru baza de date).
Prelucrarea rezultatelor si compararea in baza de date se va executa pe baza unui soft dedicat acestui tip de aplicatie, capacitatea de stocare a mijloacelor informatice fiind de un milion de profile genetice.
Nu trebuie uitat ca:
valoarea unui profil genetic si informatiile aduse de catre acesta este o continuare a calitatii muncii indeplinite de catre personalul care efectueaza cercetarea la fata locului si celui care instrumenteaza cazul.
scena infractiunii ofera de cele mai multe ori informatii pretioase despre evenimentele petrecute acolo. Aceste informatii trebuiesc valorificate.
Sa presupunem ca normele metodologice privind ridicarea, ambalarea si conservarea urmelor biologice provenite din campul infractional destinate analizei genetice, sunt respectate intocmai.
Totalitatea insusirilor si a laturilor esentiale in virtutea carora informatiile aduse de un profil genetic pot fi valorificate corespunzator se bazeaza pe o insiruire de etape care incep din momentul in care a avut loc infractiunea. Chiar de la intrarea in scena infractiunii a personalului autorizat este bine sa se tina seama ca prin necunoasterea adecvata a materiei asupra careia este orientata activitatea subiectului, pot fi introduse erori care odata inglobate in schema sunt foarte greu de corectat.
În sensul trecerii in revista a performantelor elementelor probatorii posibile pentru aceste tipuri de analize, va sugerez urmatoarea comparatie: o urma papilara lasata pe un fragment de sticla contine suficiente celule epiteliale din care se poate efectua un profil genetic, dar contine totodata si suficiente elemente pentru a o putea valorifica in sistemul AFIS. Imaginati-va ca, aceeasi urma este lasata pe o suprafata rugoasa care nu permite pastrarea unor elemente suficiente de identificare papilara. Din aceasta urma se poate extrage suficient material genetic pentru a obtine un profil ADN al persoanei care a creat-o. Pentru exemplificare, redau cateva obiecte de pe care s-a reusit valorificarea unor astfel de urme biologice in laboratorul de analize genetice al Politiei Romane: pahar, sticla, ac de seringa, bricheta, diverse alte urme de contact etc.
Nu trebuie uitat ca pentru o valorificare completa a urmelor identificate la fata locului sunt necesare si profilele de referinta ale unuia sau mai multor posibili suspecti sau o baza de date unde profilul genetic al urmei analizate trebuie cautat si identificat.
Raportul de expertiza a laboratorului contine printre alte informatii legate de natura urmelor analizate, pozitionarea lor si "tabelul alelelor" care prezinta profilul genetic pentru fiecare proba sau urma analizata.
Profilul genetic este o lista de alele a anumitor loci (markeri genetici) din genomul uman. Pentru a intelege probatoriul amprentei genetice este necesar, mai intai sa se inteleaga continutul acestui tabel. Rezultatele se obtin intai sub forma de electroferograme - figura 1- acestea sunt apoi trecute automat sub forma de numere intr-un tabel.
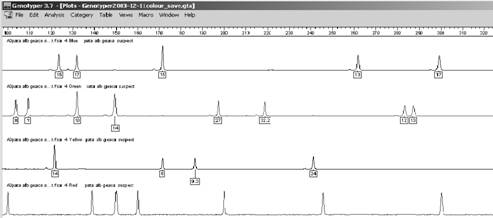
Figura 1
În tabelul de mai jos sunt prezentate profilele genetice pentru cinci probe: o urma de sange de la fata locului si patru probe de referinta recoltate de la suspecti diferiti.
|
Loci |
D3S1358 |
vWA |
D16S539 |
D2S1338 |
AMEL |
D8S1179 |
D21S11 |
D18S51 |
D19S433 |
TH01 |
FGA |
|||||||||||
|
Urma sange |
X |
Y | ||||||||||||||||||||
|
Suspect 1 |
X |
Y | ||||||||||||||||||||
|
Suspect 2 |
X |
Y | ||||||||||||||||||||
|
Suspect 3 |
X |
Y |
| |||||||||||||||||||
|
Suspect 4 |
X |
Y | ||||||||||||||||||||
Fragmentele de ADN luate in lucru din aceste urme biologice umane au fost analizate cu un instrument numit "ABI Prism 310 Genetic AnalyzerTM" produs de compania Applied Biosystems, Inc. (ABI) utilizand kit-ul de reactivi SGM+ (pe langa acesta, in cadrul laboratorului poate fi utilizat si kit-ul IDENTIFILER produs de aceeasi companie, care analizeaza un numar de 16 loci - CSF1PO, FGA, TPOX, TH01, VWA, D3S1358, D5S818, D7S820, D8S1179, D13S317, D16S539, D18S51, D21S11, D19S433, D2S1338, amelogenin).
Pentru fiecare locus sunt luati in studiu un anumit numar de alele unanim acceptate de comunitatile forensic. În mod normal, fiecare individ poseda doua alele diferite (heterozigot) sau identice (homozigot) pentru fiecare marker (locus). Aceste alele sunt etichetate cu numere in functie de anumite proprietati fizico-chimice ale fragmentelor de ADN studiate.
Variabilitatea genetica a speciei umane poate fi evidentiata prin diferentele care exista intre toti indivizii, cu exceptia gemenilor monozigoti. Culoarea parului, inaltimea, forma, toate aceste caracteristici sunt exprimarea unor alele din patrimoniul genetic.
Cu cat numarul de alele este mai mare pentru un marker ADN dat, cu atat este mai mare si numarul de genotipuri posibile. O combinatie de 10 loci, fiecare cu cate 10 alele permite existenta a 2,5x1017 genotipuri.
Pe de alta parte recombinarea este un proces prin care descendentii creeaza o combinatie de gene diferita de cea a parintilor. În timpul procesului de meioza sau de formare a gametilor, fiecare dintre celulele de reproducere primeste aleatoriu cate un cromozom din fiecare pereche, adica in total 23 de cromozomi. Sunt astfel posibile in total 223 (8,4 milioane) de combinatii cromozomiale atat in ovule cat si in spermatozoizi. Pentru celula rezultata prin unirea spermatozoidului cu ovulul sunt posibile 223 x 223 = 70 trilioane de combinatii diferite, fiecare dintre acestea ingloband cate jumatate din patrimoniul genetic al ascendentului matern si din cel al ascendentului patern.
Fiecare alela are o anumita frecventa de aparitie in cadrul speciei umane, care releva posibilitatea existentei acesteia la alti indivizi. Combinatiile alelor - doua alele diferite (heterozigot) sau identice (homozigot) - din cadrul unui locus vor avea la randul lor o alta frecventa, evident mai mica. Utilizandu-se in studiu mai multi markeri genetici, posibilitatea de regasire a aceleiasi combinatii de alele scade foarte mult apropiindu-se de unicitate.
De exemplu, puterea de discriminare a persoanelor pe baza profilului genetic obtinut cu kit-ul SGM+, utilizand in calcul frecventele alelelor studiate, este de ordinul 1:3,3X1012, iar in cazul utilizarii kit-ului Identifiler este de 1:2.1x1017.
Revenind la exemplul dat mai sus, ne punem intrebarea: care din cei patru suspecti a creat urma de sange?
În general compararea a doua sau mai multor profile genetice (genotipuri) poate conduce la formularea uneia din urmatoarele trei concluzii:
Exista identitate intre genotipurile probelor si nu exista diferente intre probe care nu pot fi explicate. În acest caz se precizeaza si semnificanta statistica a rezultatului, la care vom reveni in continuare.
Excludere - prin compararea genotipurilor probelor rezulta diferente care pot fi explicate numai prin faptul ca acestea provin din surse diferite.
Concluzie incerta in privinta rezultatelor sau a interpretarii acestora in sensul unei identificari. Aceasta poate constitui concluzia finala atunci cand doi specialisti nu ajung la un punct de vedere comun pe baza analizei si discutarii rezultatelor si daca se considera ca nu se dispun de suficiente informatii pentru a fi formulata o concluzie certa.
În cazul nostru se poate vedea clar ca ansamblu de numere in ordinea determinata de proba de sange este repetitiv cu acesta doar in cazul suspectului 3.
Numai pe baza acestui rezultat se poate spune cu certitudine ca urma de sange a fost creata de suspectul 3?
Care este sansa ca aceasta urma sa fi fost creata de o alta persoana cu profil genetic identic cu al suspectului 3?
Este posibil ca profilul suspectului 3 sa fie identic cu al urmei de sange numai datorita unei erori de genotipare?
Ca raspuns la intrebarea 3 se poate afirma de la inceput ca in sistemul de analize genetice din dotarea Politiei Romane o astfel de ipoteza este practic imposibila datorita modului de primire a probelor, etichetare a acestora si mai mult fluxul tehnologic de obtinere a profilelor genetice nu permite o incrucisare a probelor. Acest sistem inglobeaza doua linii de prelucrare si obtinere a informatiilor, una pentru urmele biologice ridicate din campul infractional - oricare ar fi acestea - iar alta in exclusivitate pentru obtinerea profilelor genetice de referinta si alimentare a bazei de date.
Pentru a raspunde la intrebarile 1 si 2 este necesar sa se aminteasca cateva notiuni de baza ale statisticii si probabilitatilor.
Prima lege a probabilitatii spune ca probabilitatea poate lua valori in intervalul 1 pana la 0 si numai un eveniment care are loc cu certitudine are probabilitatea 1. Prima lege a probabilitatii poate fi scrisa astfel:
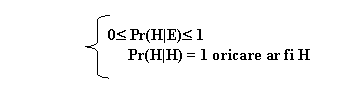
Unde Pr (H|E) - "probabilitatea lui H daca exista conditia E", in urmatoarele ipoteze:
Ø probabilitatea ca evenimentul H sa se fi intamplat;
Ø probabilitatea ca H sa aiba loc in viitor;
Ø probabilitatea ca H sa fie adevarata.
Daca stim ca H este adevarat, probabilitatea sa va fi 1.
A doua lege spune ca daca doua evenimente se exclud reciproc si dorim sa cunoastem probabilitatea ca unul sau altul sa fie adevarat atunci se aduna probabilitatile pentru fiecare. Deci, daca G si H sunt evenimente care se exclud reciproc, cu conditia E, atunci a doua lege a probabilitatii se poate scrie:
Pr(G sau H|E) = Pr(G|E) + Pr(H|E)
A treia lege demonstreaza ca daca doua evenimente pot avea loc simultan, atunci probabilitatea ca acestea sa fie adevarata se obtine prin multiplicarea probabilitatilor acestora.
Pentru doua evenimente J si K, a treia lege a probabilitatii poate fi scrisa astfel:
Pr(J si K|E) = Pr(J|E) Pr(K|J,E)
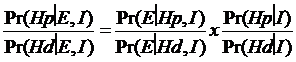
Aceasta este forma de sanse a teoremei lui Bayes care in limbajul probabilistic permite aprecierea numerica a certitudinii sau incertitudinii.
În exemplul nostru, specialistul poate anticipa ca in cazul in care fapta ajunge in instanta, acuzarea va pune in fata juratilor urmatoarea afirmatie (propozitie in sens logic):
Hp: Suspectul a creat urma de la fata locului,
iar apararea:
Hd : o alta persoana a creat urma existenta la fata locului.
În mod clar, Hp si Hd se exclud reciproc si sunt exclusive (nu mai poate fi o alta propozitie adevarata). Vom sublinia aici ca exista primul principiu al interpretarii probelor: pentru evaluarea incertitudinii oricarei propozitii este necesar sa avem in vedere cel putin o propozitie alternativa.
Sansele posterioare, care includ informatiile aduse dupa obtinerea profilului genetic identic cu al urmei de sange de la fata locului (I), pe care dorim sa le calculam se obtin prin inmultirea sanselor anterioare, fara a lua in calcul informatia de mai sus, cu un raport dintre doua probabilitati - raportul sanselor (likelihood ratio, LR):
LR = 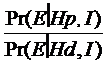
Sa presupunem, fara a intra in detalii privind modul de obtinere a rezultatului, o estimare a genotipului G in proportie de P in populatia din care face parte autorul. Deci, daca vom atribui probabilitatea P numitorului din ecuatia de mai sus, raportul sanselor devine:
LR = 1/P
Aceasta informatie transpusa practic pentru exemplul nostru devine:
P = 1/2,8 x 1014,
LR = 2,8 x 1014
ceea ce semnifica ca este de 2,8 x 1014 de ori mai probabila prima ipoteza, profilul genetic al urmei de sange de la fata locului este al suspectului 3 - implicit urma a fost creata de catre acesta - decat a doua - profilul genetic al urmei de la fata locului apartine unei alte persoane din populatia studiata, inclusiv ca urma a fost creata de catre o alta persoana necunoscuta.
Atribuirea unei valori numerice pentru numitor se face pe baza informatiilor de frecventa dispuse de catre compania producatoare a kitului de reactiv utilizat - Applied Biosystems, Inc. (ABI), in cazul nostru. Aceste valori au fost obtinute pe baza unor calcule de frecventa a alelor utilizate pentru genotipare, in diverse tipuri de populatii.
În calcul nostru frecventa (F) a acestui profil genetic in populatia caucaziana, calculata pentru kit-ul SGM+, este 3.57 x 10-12 ).
Cu alte cuvinte specialistul foloseste o estimare a proportiei din populatie care are genotipul G pentru a aprecia probabilitatea ca proba de la locul faptei sa fi fost lasata de o alta persoana.
Exprimarea rezultatului pentru cazurile simple ca cel descris mai sus utilizand exclusiv frecventa genotipului - stabilit pe baza datelor numerice ale frecventelor furnizate de producatorul kitului de reactivi utilizat - este la fel de semnificativa ca si raportul sanselor. Pentru exemplul nostru se poate afirma astfel, ca un profil genetic identic cu al suspectului 3 poate fi regasit la un numar de persoane mai mare de 3,57x1012. O reprezentare grafica a acestui rezultat este urmatoarea:
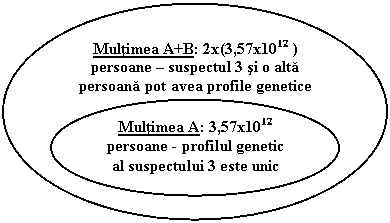
Totusi, atunci cand apar complicatii, abordarea exclusiva a frecventelor duce la concluzii gresite. Este suficient sa spunem ca rezultatele expertizei sunt prezentate in instanta, acolo unde dezbaterile nu se supun legilor stiintei sau matematicii si unde multi dintre participanti nu au nici macar cunostinte elementare de matematica. Vom constata ca sintagma "dincolo de orice dubiu rational" nu poate fi cuantificata.
Rationamentul Bayesian este cel mai bun model de care dispunem pentru interpretarea probelor stiintifice. El permite intelegerea unor aspecte altfel greu de sesizat. Desfasurarea procesului in instanta impune respectare a trei principii:
evaluarea incertitudinii oricarei propozitii se face comparativ cu examinarea a cel putin unei propozitie alternative - vor exista minimum doua puncte de vedere opuse: unul al apararii si celalalt al acuzarii. În aceasta situatie este valabila forma teoremei lui Bayes in care sunt mentionate sansele fiecarei dinte acestea si posibilitatea calcularii veridicitatii acestora, una in raport cu cealalta;
in interpretarea stiintifica a probelor exista intrebari de genul: " Care este probabilitatea datelor obtinute daca se accepta ca reala o anumita propozitie"?
interpretarea stiintifica a rezultatelor este conditionata nu numai de propozitiile opuse ci si de cadrul circumstantelor in care acestea sunt evaluate.
Abordarea cazurilor in care probele contin urme biologice provenite de la mai mult decat un singur contributor se face putin diferit. Sensibilitatea tehnicilor moderne a ajuns la un asemenea nivel incat incidenta, complexitatea si importanta unor asemenea cazuri cresc continuu. Nu este posibila abordarea in detaliu a intregii cazuistici, dar analizand un numar de asemenea situatii speram sa-l ajutam pe cititor sa dobandeasca o intelegere suficient de profunda pentru a fi capabil sa inteleaga oricare caz cu care se confrunta. Cele trei principii de interpretare a probelor ADN raman valabile si pentru aceste situatii:
evaluarea incertitudinii oricarei propozitii se face comparativ cu examinarea a cel putin unei propozitie alternative - vor exista minimum doua puncte de vedere opuse: unul al apararii si celalalt al acuzarii. În aceasta situatie este valabila forma teoremei lui Bayes in care sunt mentionate sansele fiecarei dintre acestea si posibilitatea calcularii veridicitatii acestora, una in raport cu cealalta;
in interpretarea stiintifica a probelor exista intrebari de genul: "Care este probabilitatea datelor obtinute daca se accepta ca reala o anumita propozitie"?
interpretarea stiintifica a rezultatelor este conditionata nu numai de propozitiile opuse ci si de cadrul circumstantelor in care acestea sunt evaluate.
Vom incepe cu cazul in care examinarea unei probe recoltate de la fata locului releva existenta unui amestec de urme biologice umane cu doua profile genetice distincte incluse si sunt motive sa se considere ca urmele au fost create de catre doua persoane diferite. Astfel se pot presupune urmatoarele:
alelele sunt independente atat in interiorul locilor cat si intre loci;
nu exista efecte de substructurare populationala importante din punct de vedere practic;
toti contributorii amestecurilor provin din aceeasi populatie.
Vom admite ca nu exista legatura genetica intre contributorii amestecului, iar excluderea alelelor este neglijabila din punct de vedere practic. Mai mult decat atat, vom efectua analiza ignorand intensitatile benzilor electroforetice sau inaltimile picurilor de pe electroferogramele generate de instrumentele de electroforeza, care poate sa discrimineaza contributorul majoritar de cel minoritar.
În analiza consideram doua ipoteze: ipoteza acuzarii Hp si ipoteza apararii Hd. Aceasta semnifica faptul ca putem folosi forma de calculul al sanselor, conform teoremei lui Bayes, iar informatiile aduse de analiza ADN se pot trata separat de celelalte informatiile, care sunt evaluate prin sansele anterioare (preliminare).
Interpretarea unui amestec depinde foarte mult de imprejurarile crimei. Calculele care se efectueaza trebuie sa tina cont de urmatorul aspect: regasirea ADN suspectului pe un obiect (o proba) care apartine sau este recoltata de la victima sau ADN al victimei pe obiecte (probe) ale suspectului.
Pentru interpretarea acestor rezultate sunt formulate doua ipoteze:
i. suspectul sau victima, in functie de caz, este contribuitor in amestecului de profile genetice al ADN extras din urma analizata alaturi de victima sau respectiv suspect;
ii. nu exista o legatura de origine intre amestecul de profile genetice al ADN extras din urma analizata si profilul genetic al suspectului sau respectiv victimei, cu alte cuvinte nu acesta a fost contributor la crearea urmei biologice, ci o alta persoana necunoscuta.
În urma calculelor de biostatistica aferente si al raportului dintre ipoteza i. si ii. se concluzioneaza printr-o valoare numerica varianta posibila.
Pentru o mai buna intelegere a necesitatii formularii acestor ipoteze, precum si a calculelor biostatistice efectuate, vom trece in revista cele mai frecvente cazuri utilizand un singur locus.
În primul exemplu consideram un caz in care avem argumente intemeiate pentru a crede ca ADN-ul victimei este prezent in proba, cum ar fi de exemplu o proba vaginala ridicata intr-un caz de viol (ignoram posibilitatea unei extractii diferentiale a spermei si celulelor epiteliale vaginale care are menirea de a separa fractia masculina de cea feminina si de a obtine separat profilele genetice ale celor doua fractii). În acest caz proba ridicata va contine ADN de la victima si de la agresor si vom gasi atat alelele victimei cat si cele ale suspectului. Vom presupune totodata faptul ca victima a declarat ca nici un alt ADN barbatesc nu poate fi prezent in proba.
Alele luate in calcul sunt 15, 16, 17 si 18 din cadrul locusului D3S1358. presupunem ca alela 15 si 18 provine de la suspect, iar 16 si 17 de la victima - Figura 2.
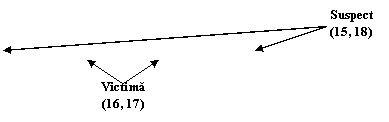
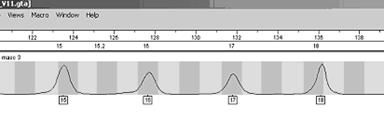
Figura 2. Amestec de 4 alele
Proba in litigiu are 4 alele pentru acest locus, doua dintre acestea fiind identice cu ale victimei iar doua identice cu cele ale suspectului. Propozitia acuzarii este urmatoarea:
Hp: Proba in litigiu contine ADN de la victima si de la suspect.
Propozitia apararii poate fi de tipul:
Hd: Proba ridicata de la fata locului contine ADN de la victima si de la o persoana necunoscuta.
Vom nota cu Ec profilul amestecului si cu Gv si Gs genotipurile victimei si respectiv suspectului. Raportul sanselor (LR) pentru acest amestec este:
LR = 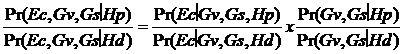
![]()
Întrucat nimic din ipotezele Hp sau Hd nu poate afecta incertitudinea referitoare la Gv si Gs, al doilea raport de mai sus este 1 si deci
LR = 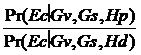
![]()
Gs nu influenteaza incertitudinea noastra cu privire la autor, deci:
LR = 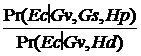
Numaratorul este 1 intrucat, daca Hp este adevarata, profilul probei in litigiu este exact cel asteptat. Numitorul reprezinta probabilitatea ca o persoana necunoscuta, nelegata genetic de victima sa contribuie cu alelele 15 si 18. Întrucat aceasta probabilitate nu depinde de genotipul suspectului, valoarea ei este, conform teoremei Hardy Weinberg 2 (p15p18), unde p15 si p18 sunt probabilitatile de regasire ale alelor 15 si 18 in populatie. În consecinta:
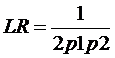
Acest rezultat stabileste modul uzual de "scadere" a genotipului victimei din genotipul probei.
Daca cei doi contributori ai amestecului au o alela in comun, profilul genetic al ADN extras din urma biologica analizata va contine numai trei alele. Putem distinge cazurile in care victima este homozigot sau este heterozigot. Pentru primul caz sa luam locusul THO1 cu alelele 6, 7 si 9,3 - Figura 3.


Figura 3 - amestecul de trei alele, victima homozigot
Daca victima este homozigot (9.3, 9.3) (Figura 2), un rationament similar cu cel din exemplul cu patru alele duce la acelasi rezultat:
LR = 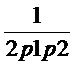
Daca victima este heterozigot (13, 14), iar suspectul este homozigot (12, 12) (Figura 4), numaratorul raportului sanselor ramane egal cu 1, dar evaluarea numitorului necesita calcule suplimentare. Este necesar sa luam in considerare trei posibilitati pentru genotipul G al persoanei necunoscute: (12, 12); (12, 13) si (12, 14) - Tabelul 2.

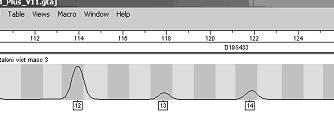
Figura 4. Amestec de trei alele - victima heterozigot.
|
i |
Gi |
Tabelul 2 - posibilitati pentru genotipul persoanei necunoscute
Pentru evaluarea numitorului din ecuatia care defineste raportul sanselor, se aplica legea probabilitatii totale:
Pr(EcïGv,Hd) =
= ![]()
Deoarece am ignorat diferentele de intensitate ale picurilor, oricare dintre Gi plus genotipul victimei vor conduce la profilul probei in litigiu, deci:
Pr(EcïGv,Gi,Hd) = 1, i = 1, 2, 3
si
Pr(EcïGv,Hd) = 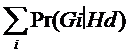
Probabilitatile Gi nu depind de Hd, ele sunt date de produsele frecventelor alelelor 12, 13 si 14 (p1, p2 si respectiv p3). Raportul sanselor devine astfel:
LR = 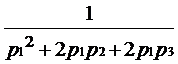
Se observa faptul ca raportul sanselor este mai mic decat valoarea 1/pi2, corespunzatoare cazului in care suspectul ar fi fost contributor unic al unei urme cu genotipul (12, 12). Prezenta in urme a genotipului victimei are ca efect scaderea probabilitatii ipotezei acuzarii, deoarece a crescut numarul de ipoteze posibile, pentru explicarea datelor aduse de catre profilul genetic in cazul in care Hd este adevarata.
Uneori probele de la fata locului pot contine ADN provenind de la mai mult decat o singura persoana, dar numai una cunoscuta este suspectata ca fiind contributoare. În acest caz trebuie calculata probabilitatea de regasire a profilului genetic cunoscut, in acest amestec.
Daca luam cazul locusului TH01 care prezinta pentru un amestec patru alele (6, 7, 9, si 9.3) care includ si genotipul unui suspect unic (7, 9) - Figura 5, se pot formula urmatoarele ipoteze:
Hp: Proba in litigiu contine ADN provenind de la suspect si de la o persoana necunoscuta.
Hd: Proba in litigiu contine ADN de la doua persoane necunoscute.
Folosind acelasi rationament ca si in cazul in care victima este contributoare, se poate demonstra ca:
LR = 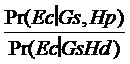
Genotipurile Gi, posibil componente ale probei in litigiu sunt urmatoarele:
|
i |
Gi |
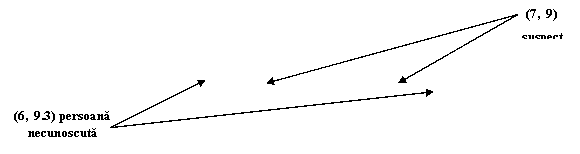
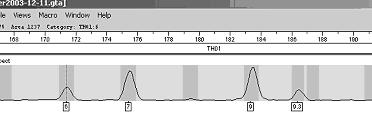
Figura Amestec cu patru alele
Observam faptul ca G1 = Gs. Legea probabilitatii totale ne da:
Pr (EcïGs,Hp) = 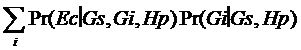
unde Gi este genotipul contributorului necunoscut al amestecului, care exista pe langa genotipul suspectului.
În urma altor calcule de probabilitate, pornind de la premisa ca cei doi contributori nu sunt inruditi genetic, astfel incat Gi si Gj sunt independente, ignorand diferentele de intensitate ale picurilor electroferogramei si atribuind valoarea 1 pentru toate cele 6 combinatii Pr(EcïGi,Gj,Hd) = 1, raportul sanselor devine:
LR = 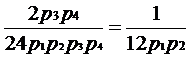
unde p1 = p7, p2 = p9
p3 = p6 ;I p4 = p9.3
Raportul sanselor este micsorat de sase ori in comparatie cu valoarea pe care ar fi avut-o in situatia in care genotipul suspectului (7; 9) ar fi fost inclus intr-o singura urma biologica, cu profilul 7, 9. Daca alelele 7, 9 sunt prezente in mod obisnuit in populatie (de ex. p7 = p9 = 0,3, ceea ce da 12p7p9 > 1) raportul sanselor este de fapt mai mic decat 1, aceasta valoare semnificand faptul ca este mai probabil ca o urma cu profilul 6, 7, 9, 9.3 sa provina de la persoane necunoscute decat sa provina de la suspect si de la o persoana necunoscuta. Deci rezultatele sunt in favoarea apararii, ceea ce demonstreaza ca este mai important sa se interpreteze rezultatele genotiparii decat sa se faca afirmatii de tipul "o persoana oarecare nu este exclus ca posibil contributor".
Atunci cand suspectul este heterozigot, de exemplu alele (15, 17) din cadrul locusului vWA, iar proba de la fata locului prezinta trei alele 15, 17 si 18 - Figura 6, raportul sanselor este:
LR = 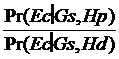
iar genotipurile care pot fi componente ale profilului probei de la fata locului sunt:
|
i |
Gi |
Pr(GiïHp) |
|
2p15p17 |
||
|
2p15p19 |
||
|
2p17p18 |
||
|
P152 |
||
|
P172 |
||
|
P182 |
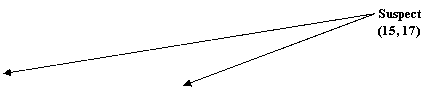
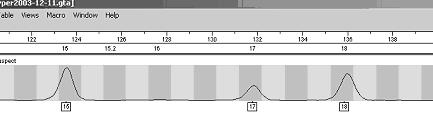
Figura 6. Amestec de trei alele, suspectul heterozigot (15, 17)
Daca G1 = Gs, atunci numaratorul raportului sanselor LR devine:
Pr(EcïGs,Hp) =
= ![]()
dar numai variantele i = 2, 3, 6 pot fi ale unui profil genetic al ADN extras din proba de la fata locului (15, 17 si 18) si pentru aceste valori Pr(EcïG1,Gi,Hp) = 1.
Adunand termenii sumei se obtine:
Pr(EcïGs,Hp) = 2p15p18 + 2p17p18 +p182
Existand 12 combinatii a celor doua genotipuri, care formeaza un profil identic cu cel al probei de la fata locului, se simplifica numitorul raportului sanselor, obtinandu-se relatia:
Pr(EcïGs,Hp) = 12p15p17p18(p15+p17+p18)
Dupa care raportul devine:
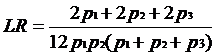
unde p1 = p15, p2 = p17 si p3 = p18.
Exista situatii in care genotipurile a doi suspecti pot fi incluse intr-un amestec de profile genetice al ADN extras dintr-o proba. Pentru acest exemplu presupunem ca exista date conform carora crima a fost comisa de doi autori. Un asemenea exemplu este un viol dublu, unde proba in litigiu este fractiunea masculina extrasa de pe un tampon cu care s-a recoltat secretia vaginala de la victima. Doi suspecti au fost arestati pentru comiterea crimei, pentru motive pe care le includem in probele non-ADN, I. Sa presupunem ca cei doi suspecti sunt judecati impreuna, caz in care propozitia acuzarii poate fi formulata astfel:
Hp: Proba in litigiu contine ADN de la cei doi suspecti.
Totusi, in privinta apararii ar fi nerealist sa se anticipeze o singura propozitie, chiar si numai pentru faptul ca cei doi suspecti cel mai probabil sunt reprezentati de doi aparatori. Putem presupune ca apararea va formula urmatoarele ipoteze:
Hd1: Proba in litigiu contine ADN provenind de la suspectul 1 si de la o persoana necunoscuta.
Hd2: Proba in litigiu contine ADN provenind de la suspectul 2 si de la o persoana necunoscuta.
Hd3: Proba in litigiu contine ADN provenind de la doua persoane necunoscute.
În Figura 7 este prezentat un exemplu de amestec de profile genetice pentru locusul TH01 in care sunt incluse genotipurile a doi suspecti heterozigoti:
- suspect 1: (9.3, 11);
- suspect 2: (10, 13.3).
Pe baza rezultatelor obtinute anterior si stipuland premisele privind independenta profilelor, obtinem:
Pr(EcïHp) = 1
Pr(EcïHd1) = 2p10p13.3
Pr(EcïHd2) = 2p9.3p11
Pr(EcïHd3) =24 p9.3p11 p10p13.3
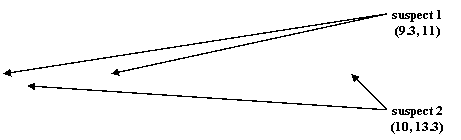
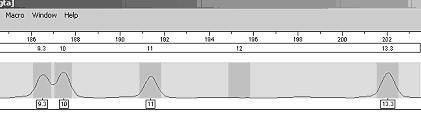
Figura 7. Electroferograma pentru amestecul cu patru alele
obtinut pentru locusul TH01
Cand exista mai mult de doua ipoteze, este necesar sa folosim forma generala a teoremei lui Bayes pentru a calcula probabilitatile ulterioare. Folosind legea probabilitatii totale, obtinem pentru propozitia acuzarii:
Pr(HpïEc) =
= 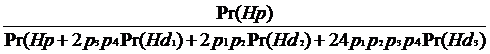
În cazul nostru: p1 = p9.3, p2 = p11, P3 = p10, P4 = p13.3
O solutie a acestei probleme este de a se admite o pereche de ipoteze alternative si sa se efectueze calculele pentru fiecare din cele doua ipoteze. Se ajunge astfel la un raport al sanselor separat. Rezultatele se pot reprezenta in urmatorul tabel:
|
Numitor |
Numarator |
||
|
Hp |
Hd1 |
Hd2 |
|
|
Hd1 |
1/(2p9.3p11) | ||
|
Hd2 |
1/(2p9.3p11) |
p3p4/( p9.3p11) | |
|
Hd3 |
1/(242p9.3p11p10p13.3) |
1/(12 p9.3p11) |
1/(12 p10p13.3) |
Desigur, tabelul se poate simplifica in situatia in care unul dintre suspecti pledeaza vinovat.
Pentru cazurile discutate am admis, atat pentru ipoteza Hp cat si pentru Hd ca ADN-ul victimei este regasit in proba in litigiu. Pot fi totusi situatii in care aceasta prezumtie nu este valabila. Proba in cazul unui viol poate fi amestec de sange si sperma de pe un obiect de pe pat. Daca profilul genetic al probei include genotipurile victimei si suspectului, iar victima identifica obiectul respectiv, propozitia acuzarii va fi similara cu cea de mai sus:
Hp: Proba in litigiu contine ADN provenind de la victima si de la suspect.
În acest caz pentru propozitia apararii pot fi admise urmatoarele variante:
Hd1: Proba in litigiu contine ADN provenind de la victima si de la o persoana necunoscuta.
Hd2: Proba in litigiu contine ADN provenind de la suspect si de la o persoana necunoscuta.
Hd3: Proba in litigiu contine ADN provenind de la doua persoane necunoscute.
Raportul sanselor poate fi folosit analizand simultan doua ipoteze:
urma contine ADN provenind de la victima si de la o persoana necunoscuta.
urma contine ADN provenind de la doua persoane necunoscute.
Evaluarea raportului sanselor se face ca in cazurile de mai sus, astfel ca specialistul va avea posibilitatea sa concluzioneze: " Probele pledeaza pentru faptul ca este de "LR" ori mai probabil ca prima dintre aceste ipoteze sa fie adevarata decat ca cea de-a doua ipoteza."
Stabilirea faptului ca victima a fost sau nu violata pe obiectul respectiv pe care exista urma depinde nu numai de probele ADN ci si de alte circumstante pe care instanta le va lua in considerare. Daca instanta va stabili ca, intr-adevar obiectul de pe pat este asociat cu violul, atunci este util sa se analizeze urmatoarele ipoteze:
urma contine ADN provenind de la victima si de la suspect;
urma contine ADN provenind de la victima si de la o persoana necunoscuta.
Evaluarea raportului sanselor pentru aceasta alternativa decurge ca mai inainte. Concluzia specialistului va fi de forma: "Probele ADN demonstreaza faptul ca este de LR ori mai probabil ca prima dintre aceste propozitii sa fie adevarata."
Interpretarea amestecurilor se poate face numai in contextul rapoartelor sanselor (LR). Spre deosebire de urmele cu un singur contributor, in cazul amestecurilor de profile genetice nu este certa nici una dintre cele doua ipoteze, de aceea se face raportul celor doua probabilitati, raportul sanselor. Prezentarea probabilitatii unei singure propozitii poate induce in eroare.
În exemplele prezentate au fost luate in calcul un singur locus din cei 12 (kit-ul SGM+) sau 16 (kit-ul Identifiler). Utilizarea intregului sistem de loci face posibila discrimare intre ipotezele acuzarii si apararii cu o valoare numerica care poate fi numita certitudine.
Am ignorat in explicatiile date orice informatie, cum ar fi intensitatea (aria) sau inaltimea picurilor care ar putea fi un indicator al cantitatilor relative de ADN ce provine de la un contributor din cadrul unui amestec. Luand in calcul diferentele de intensitate dintre picurile unei electroferograme ce contine un amestec de doua profile genetice, se poate creste capacitatea de discriminare a analizei.
Exista de asemenea si alte metode care pot stabili anumite combinatii genotipice care in functie de matodele de interpretare pot furniza informatii adiacente cu o probalilitate aproape de certitudine.
|
Politica de confidentialitate |
| Copyright ©
2024 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |
