Genetica - stiinta a ereditatii, variabilitatii si reproducerii organismelor, este privita cu interes stiintific de elevi, studenti, cercetatori, medici etc. Ea reprezinta o sinteza unitara si creatoare a principalelor descoperiri ale citologiei, fizicii, chimiei, matematicii si ciberneticii. Multitudinea descoperirilor genetice au medernizat stiinta despre viata si au impus-o, prefigurANDu-i un nou viitor.
Evolutia fizico-chimica a materei pe Terra a condos la cresterea gradului de complicare structurala, ceea ce a constituit premiza, aparitiei vietii pe pamant, punct de plecare a evolutiei biologice.
Primului organism viu ii era caracteristica autoconservarea si autoreproducerea, insusiri care il deosebeau net de lumea nevie. Mentinerea structurilor caracteristice viului se realizau cu consum de energie, preluata direct din mediu, din energia solara. Stocarea si utilizarea acestei energii se realizeaza printr-o alta insusire caracteristica doar organismelor vii - metabolismul.
Carcateristicile viului puteau fi mentinute si accentuatein generatii successive, decat prin pastrarea si perfectionarea structurilor ce le0au facilitat aparitia. Era deci absolut necasara existenta unor substante capabile sa detina si sa transmita aceste instructiuni de la un organism viu la organismele descendente; era deci necesara existenta unui material genetic. Exista substante forte simple, asa cum sun virusurile, ce detin in structura lor 2 tipuri de substante organice - proteine si acizi nucleici .
Acidul ribonucleic (ARN) - material genetic pentru unii virusi
Unii virusi maturi sau virioni au in compozitia lor numai acizi nucleici si proteine. Proprietatea de pastrare si transmitere a informatiei geneticen e determinate experimental prin infectia frunzelor de tutun cu VMT (virusul mozaicului tutunului)- fig.1.
Particula virotica este
alcatuita dintr-o macromalecula de ARN si un manson proteic. Studiile
electromicroscopice arata ca miezul particulei este
alcatuit din ARN monocatenar, dispus spiralat partea exterioara - ADN-nul
proteic - monomere. Aranjarea monomerilor este facuta
dupa un plan spiral ce cuprinde 130 de spire. Un
monomer proteic cuprinde 158 molecule de aminoacizi incatenati intr-o secventa
specifica fiecarei suse de VMT. Cuprinde un lant
monocatenar ce contine 6400 de nucleotide, dispuse intr-o succesiune
caracteristica pentru fiecare
Infectiozitate si autoreproducere VMT. VMT nu prezinta un mecanism propriu de infectare a celulei gazda. El patrunde in celulele frunzelor de tutun printr-o leziuzne cauzata in special de intepaturile insectelor. Patrunsa in celula gazda, particular virotica se elibereaza de mansonul proteic. Dupa cateva ore - perioada de incubatie ARN-ul virotic patrunde in nucleul celulei gazda si isi initiaza autoreproducerea. Acest proces se realizeaza pe seama aparatului metabolic al celulei gazda. Adica macromolecula de ARN si VMT dirijeaza intregul metabolism al celulei gazda spre biosinteza de noi macromolecule de ARN virotic. Apoi, noile macromolecule de ARN virotic determina biosinteza monomerilor, respective a mansoanelor proteice.
ARN
 Fig.1
Fig.1
Reconstituirea artificiala a VMT prin hibridari moleculare . Particula virotica a VMT a fost dezagregata in cele 2 parti componente. Astfel sub actiunbea acidului acetic se pot extrage numai proteinele native iar cu ajutorul fenolului (acid carbolic) s-a extras ARN, particulele proteice fiind denaturate.
Proteinele virotice nu sunt capabile sa indica nici boala si nici isnteza de proteine virale., In schimb ARN-ul viral a determinat sinteza a sute de noi particule virale ce detineau ARN-ul si proteinele caracteristice, ceea ce a dus la concluzia ca ARN este materialul genetic al VMT.
Acidul dezoxiribonucleic - ADN - material genetic pentru alta categorie de virusi
Virusii ADN, care contin numai ADN si proteine caracteristice., componente ce pot fi separate prin acelasi tratament cu acid acetic si fenol. Cele 2 componente pot fi introduse separate in celula gazda. Pentru astfel de ecperimente a fost utilizat un bacteriofag cu structura foarte simpla numit phi X Acesta are drept gazda bacteria Esherichia coli . Ca orice bacterie tipica aceasta are un perete celular alcauit din polizaharite, care se opune penetrarii de macromolecule. Din aceasta cauza peretele celular trebuie indepartat. O astfel de bacterie , fara perete cellular, poarte denumirea de sfenoplast.Daca dintr-o cultura de sfenoplasti este introdus And virusului phi C 174, dupa cateva ore, apar numerosi virusi. Daca intr-o alta cultura de sfenoplasti se introduce proteina virusului, nu se vor obtine nici virusi si nici proteine virusale. De aici concluzia ca , ADN reprezinta materialul genetic al virusului phi X 174.
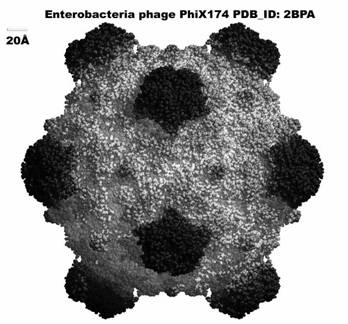
Alti virusi ADN , ce au drept gazda tot E. coli prezinta o structura mult mai complexa decat phi X 174. in aceasta categorie intra 7 tipuri de fagi. Acestia au in esenta, doua parti distincte: capul fagului - cu forma sferica sau poliedridca si coada fagului
Capul fagului prezinta la exterior un invelis proteic alcatuit din monomere proteice - capsomere. Acestea formeaza capsoda virala in interiorul careia se afla o uriasa molecula de ADN.
Coada fagului se leaga de capsida printr-un disc intermediary. Coada are forma cilindirca, alcatuita din proteine contractile. In lumenul partii contractile a cozii se gaseste un ax codal tot de natura proteica.In partea distala coada prezinta rasucite in jurul ei sase fibrile proteice numite fibrile codale, care au rolul de a fixa bacteriofagul de peretele cellular al bacteriei. La extremitatea cozii se gaseste placa bazala, de obicei, de forma hexagonala, ce produce lizozimul fagic cu rol de distrugere al peretelui celular bacterian.
Infectiozitatea. LA acest tip de bacteriofagi infectia este activa. Ea se desfasoara in cateva etape successive.
In prima etapa se produce o adsorbtie reversibila a fagului de paretele bacterian ;
Sub actiunea unor substante din peretele bacterian, fibrilele cozii se destind absorbindu-seputernic si ireversibil in jurul zonei de infectare - adsorbtie ireversibila ;
Sub actiunea lizozimului fagic, se produce o bresa fizico-chimic in peretele bacterian si zona contractila a cozii se contracta - faza contractila a infectiei fagice ;
Ca urmare a contractiei axul codal este expulzat, Patrunzand in interiorul bacteriei, el antreneaza si macromolecalu de ARN ; aceasta este faza infectarii ;
Macromolecula de ADN fagic este transferata integral in interiorul bacteriei. La exteriorul bacteriei ramane particular fagica, lipsita total de acid nucleic, asa numita fantoma fagica ;
Aceasta dovedeste ca materialul ereditar al fagului este reprezentat de ADn, nu si de proteine. Intregul process al infectiei fagice dureaza aproximativ un minut.
Recent s-areusit transferul de sfenoplasti bacterieni a ADN -ului fagului T4 in stare pura, obtinadu-se particule fagice complete. Aceasta vine sa demonstreze odata in plus ca ADN-ul este purtatorul material al ereditatii la aceasta categorie de bacteriofagi.
Atat ARN-ul cat si ADN-ul sunt prezente in toate celulele procariote si eucariote. In organismele ce contin atat ARN cat si |ADN materialul genetic este reprezentat intotdeauna de AND.
ADN-ul din cromozomul organismului procariot, AND-ul cromozomilor nucleici ai eucariotelor, cat si cromozomii organitelor celulare ale eucariotelor , materialul genetic este reprezentat intotdeuna de ADN.
ADN aflat in organitele celulare (mitocondrii, cloroplaste), constituie material genetic si au proprietatea de reproducere, detinere si transmitere de informatie genetica.
ADN-ul a fost depistat in diverse structuri ale celulei eucariote, dar cu preponderenta in cromozomii nucleari. ARN-ul depaseste cantitatea de ADN cellular, fiind situat in nucleu, citoplasma si alte organite celulare, ion spoecial in ribozomi. In acest caz ARN nu este material genetic, neavand capacitatea de autoreplicare, capacitatea pe care o detina, in cazul celulelor eucariote, numai ADN,
Formatiuni celulare implicate in realizarea mecanismelor ereditare
Modul de functionare si transmitere a informatiei ereditare depinde de structura si functiile celulei.
Componentele nucleului sunt : membrana, citoplasma, cu unul si mai multi nucleoli. Carioplasma este alcatuit din cariolimfa (sucul nuclear) si cromatina nucleara. Din cromatina nucleara, in cursul diviziunii se vor diferentia cormozimomii, care reprezinta principalul sediu al genelor, respective al informatiei ereditare.
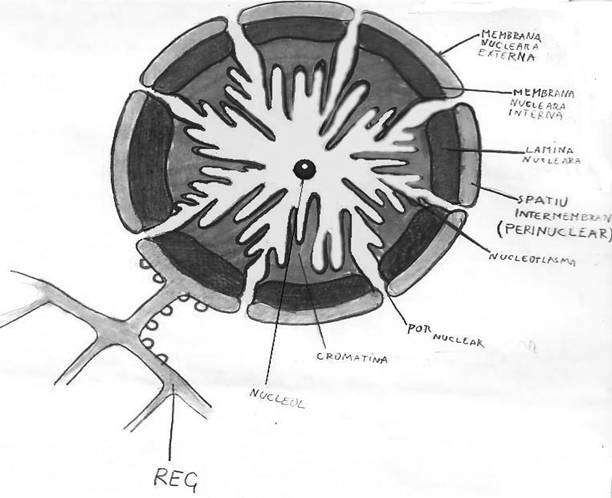
Nucleolii se constituie pe anumiti cromozomi, in regiunea ce detine informatia genetica pennru sinteza de ARN ribozomal. Aceste zone poarta denumeria de organizatori nucleolari.
In zona nucleolului ADN-ul cromozomial ramane partial despiralizat pana in metafaza constituind strangulatii secundare ale cromozomilor nucleolari. Genele ADN, ce detin informatia genetica pentru sinteza ARN-ului ribosomal, se multiplica prin replicari repetate, constituind segmente de ADN extracromozomial. Aceste segmente de ADN sunt inconjurate in interiorul nucleului de cantitati mari de ARN ribosomal, care se afla in diverse stadii de maturare. In structura nucleolului se gasesc marin cantitati de proteine. Aceste proteine, ajunse in nucleu din citoplasma sunt heterogene ca structura si functii. O parte constituie enzime implicate in sinteza ARN-ului, o alta parte reprezinta proteine ce intra in constitutia protoribozomilor nucleolari. Acestea vor trece ulterior in citoplasma, unde vor constitui ribozomii citoplasmici.
Sucul nuclear sau carioplasma, reprezinta mediul in care se realizeaza oate transformarile din nucleu. In cadrul acestui component nuclear este descrisa o matrice nucleara formata din proteine sau greutate moleculara mare, care asigura forma nucleului jucAND un rol de carioschelet si fractiune labila in care se gasesc o serie de proteine solubile cu greutate moleculara mica, ioni, ATP si apa.
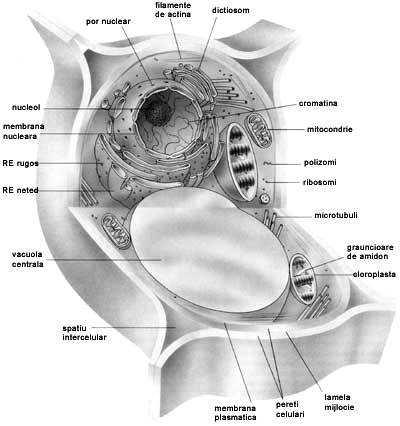
 Citoplasma
este alcatuita din protoplasma si organite
Citoplasma
este alcatuita din protoplasma si organite
 Matricea protoplasmei este partea protoplasmei necuprinsa in organitele
celulare. Prezinta un adevarat mediu
intern al celulei
contin ADN in general, toti componentii care se pot gasi intr-un organism cu
exceptia ADN. In interiorul ei se desfasoara procese
esentiale pentru celula asa cum sunt procesele metabolice ale zaharurilor,
lipidelor, proteinelor si nucleotidelor, realizarea echilibruluiionic, etc.
Matricea protoplasmei este partea protoplasmei necuprinsa in organitele
celulare. Prezinta un adevarat mediu
intern al celulei
contin ADN in general, toti componentii care se pot gasi intr-un organism cu
exceptia ADN. In interiorul ei se desfasoara procese
esentiale pentru celula asa cum sunt procesele metabolice ale zaharurilor,
lipidelor, proteinelor si nucleotidelor, realizarea echilibruluiionic, etc.
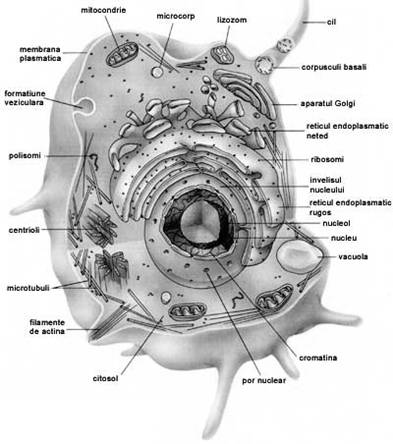
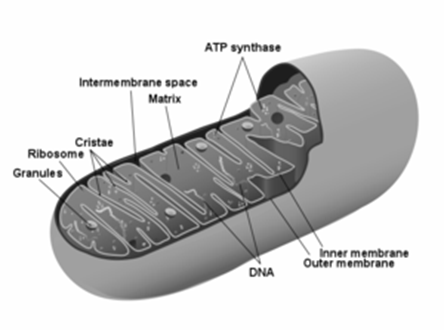 Mitocondriile sunt organite granulare sau filamentoase.
Ca structura au un invelis dublu lipopreoteic,
contribuind la realizarea celei mai mari parti a ATP-ului cellular. Din aceasta
cauza mitocondriile sunt considerate centrele energetice ale celulelor.
Mitocondriile contin ADN mitocondrial si au capacitate de autoreplicare. Acest ADN
are structura circulara, asemanandu-se cu ADN-ul bacterian. Mitocondriile detin
ribozomi mitocondriali, ARN informational, ARN de transport, precum
si enzimele necesare sintezei proteice proprii.
Mitocondriile sunt organite granulare sau filamentoase.
Ca structura au un invelis dublu lipopreoteic,
contribuind la realizarea celei mai mari parti a ATP-ului cellular. Din aceasta
cauza mitocondriile sunt considerate centrele energetice ale celulelor.
Mitocondriile contin ADN mitocondrial si au capacitate de autoreplicare. Acest ADN
are structura circulara, asemanandu-se cu ADN-ul bacterian. Mitocondriile detin
ribozomi mitocondriali, ARN informational, ARN de transport, precum
si enzimele necesare sintezei proteice proprii.
 Plastidele sunt organite caracteristice numai celulelor
vegetale. Contin pigmenti asimilatoriclorifieni, se
numesc cloroplaste. Din punct de
vedere structural, plastide;e au o membrana dubla
lipo-proteica. Platidele detin material genetic (ADN sub form circulara,
asemanatoare tipului bacterian), ribozomi liberi, ARN si intreaga gama de enzime care le
permitesa realizeze sinteze proteice. Aceste structuri le
confera plastidelor capacitatea de reproducere.
Plastidele sunt organite caracteristice numai celulelor
vegetale. Contin pigmenti asimilatoriclorifieni, se
numesc cloroplaste. Din punct de
vedere structural, plastide;e au o membrana dubla
lipo-proteica. Platidele detin material genetic (ADN sub form circulara,
asemanatoare tipului bacterian), ribozomi liberi, ARN si intreaga gama de enzime care le
permitesa realizeze sinteze proteice. Aceste structuri le
confera plastidelor capacitatea de reproducere.
Centriolii ca si
mitocondriile si plastidele , au capacitate de
autoreproducere, detinand material genetic ADN. Sunt organite caracteristice
celulelor animale, dar se pot intalni si la unele plante inferioare de tipul algelor
unicelulare. In etapa de sinteza din interfaza  celulara, cele doua manunchiuri de tubusoare se separa si fiecare determina
sinteza unui nou manunchi dispus perpendicular pe cel vechi. Prin organizarea
fusului de diviziune, asigura deplasarea cromozomilor, iar la celulele cu
miscare amoeboidale, tot centriolul este coordonatorul
acestor deplasari.
celulara, cele doua manunchiuri de tubusoare se separa si fiecare determina
sinteza unui nou manunchi dispus perpendicular pe cel vechi. Prin organizarea
fusului de diviziune, asigura deplasarea cromozomilor, iar la celulele cu
miscare amoeboidale, tot centriolul este coordonatorul
acestor deplasari.
 Ribozomii nu au capacitate
autosintetica, nu contin material genetic. Ribozomi incep sa
se sintetizeze in nucleu la nivelul nucleolilor sub forma de protoribozomi sau
ribozomi nucelolari, care trec apoi in citoplasma, unde li se adauga un nou set
de proteine, care le confera proprietati specifice.
Ribozomii nu au capacitate
autosintetica, nu contin material genetic. Ribozomi incep sa
se sintetizeze in nucleu la nivelul nucleolilor sub forma de protoribozomi sau
ribozomi nucelolari, care trec apoi in citoplasma, unde li se adauga un nou set
de proteine, care le confera proprietati specifice.
ORGANIZAREA STRUCTURALA A ACIZILOR NUCLEICI SI DISPUNEREA ACIZILOR NUCLEICI IN CROMOZOMII DIVERSELOR TIPURI DE ORGANISME
In alcatuirea acizilor nucleici participa trei substante chimice : baze azotate, glucide de tip pentozic si radicalii ai acidului fosforic.
Bazele azotate nsunt de doua tipuri:
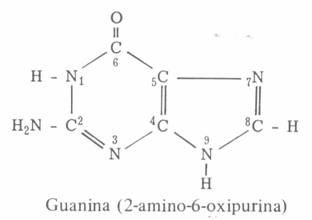
Bazele purinice sunt identice la ARN si ADN - adenina si guanina, pe cand cele pirimidice la ADN sunt reprezentate de citozina si timina, iar la ARN in locul timinei intra uracilul.
Pentozele se prezinta sub forma ciclica furazonica (heterociclu pentagonal. In componenta ARN-ului intra riboza, air ADN-ul se formeaza cu dezoxiriboza.
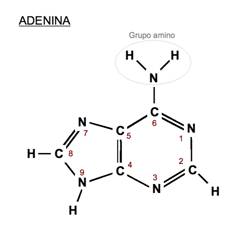
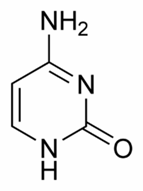
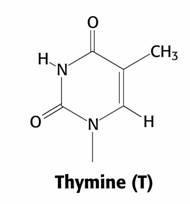
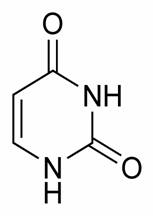
Citozina Uracil
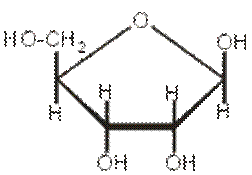
Pentozele care intra in componenta acizilor nucleici sunt riboza si dezoxiriboza
O
![]() OH
OH

 CH2 OH
CH2 OH
H H
![]()
![]() OH H
OH H
![]()
OH H
RIBOZA DEZOXIRIBOZA
Baza azotata legata de pentozaprin lagatura azola C1 , alcatuieste o nucleozida. Prin esterificarea nucleozidei cu acid ortofosforic in pozitia C5 a pentozei se formeaza o nucleotida.
In cadrul macromoleculelor de acizi nucleici incatenarea nucleotidelor se realizeaza prin esterificarea cu acid ortofosforic la C3 al pentozelor.
STRUCTURA MOLECULARA A ADN
Desi acizii nucleici au fost identificati inca din anul 1869 de catre F Miescher si izolati in 1899 de R altmann, prima ipoteza asupara alcatuirii lor moleculare a fost formulata abia in anul 1931 de catre Levene (ipoteza tetranucleotidica). Structura moleculara reala a acizilor nucleici, in primul rand al ADN-ului, a fost stabilita de catre Watson si Crick, care au alcatuit un model potrivit caruia ADN-ul nativ poseda o structura primara, monocatenara si una secundara, bicatenara.
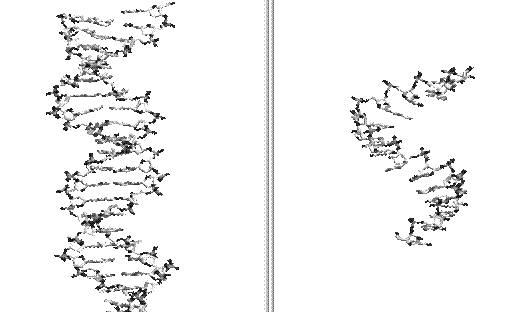
ADN bicatenar ADN monocatenar
Structura primara monocatenara a ADN-ului rezulta ca urmare a esterificarii monodezoxi-ribonucleotidelor 5 cu acid fosforic la C3 al pentozei si se prezinta sub forma unui ,lant molecular lung glucido-fosforic.
Complexitatea structurii primare a ADN este data de natura secventei bazelor azotate, deoarece unitatile glucido-fosforice sunt identice si se succed invariabil.
Fiecare oraganuism viu poseda un ADN cu secventionalitate a bazelor azotate caracteristica lui. Cu cat dou organisme sunt mai indepartate genetic, cu atat diferentele de secventionalitate bazelor azotate din moleculele lorde ADN sunt tot mai mari. Tocmai aceasta distributie specifica a bazelor azotate are un rol essential si o semnificatie determinate de functionare a ADN ca substrat material al ereditatii.
Structura secundara bicatenara a ADN-ului este starea in care se gaseste in mod obisnuit, molecula de ADN, la organismele vii.
Macromolecula bicatenara de ADN este alcatuita din doua monocatene complementare. Legaturile dintre cele 2 catene se datoresc puntilor de hydrogen care se realizeaza intre bazele azotate complementare ale celor 2 catene. Aceasta complementaritate se realizeaza intre o baza purinica si una pirimidica; dar nu oricom intotdeauna adenina se leaga de timina prin doua punti de H, iar guanina de citozina prin 3 punti de H. Legaturile de H, de natura electrostatica, sunt mai labile decat celelalte legaturi chimice care realizeaza structura primara a ADN.
Legaturile de H sunt slabe dar numarul lor foarte mare realizeaza o coeziune suficienta intre catenele complementare asigurand stabilirea ADN.
Greutatea moleculara a ADN variaza de la o specie la alta. Astfel, ADN-ul fagului T2 are o greutate moleculara de 130 milioane , iar macromolecula de ADN de E.coli are o greutate moleculara de aproximativ 2 miliaarde .
Stabilitatea ADN-ului bicatenar este asigurata si de faptul ca cele 2 monocatene se rasu cesc una in jurul celeilalte, formand o dubla spirala (dublu helix). Datorita acestei rasuciri a catenelor complementare structura macromoleculei de ADN este cunoscuta sub denumirea debicatenara plectonemicala. Plectonemicalitatea este o rezultanta a complementaritatii catenelor, aceasta stare reprezentatnd configuratia optima pentru asigurarea celei mai bune stabilitati a structurii bicatenare a ADN.
STRUCTURA MOLECULARA A ARN
In lumea vie sunt cunoscute 4 tipuri de ARN si anume : ARN viral, ARN mesager, Arn ribosomal si ARN de transport.
Strucutura ARN viral reprezinta o serie de virusi, materialul lor genetic. Aceste ARNv are o structura monocatenara, cu acelasi tip de organizare ca si Arn monocatenar, cu precizarile deja cunoscute, ca pentoza este reprezentata de riboza, iar in cadrul bazelor pirimidice in locul timinei intra uracilul. La un virus ARN este reprezentat de o singura macromolecula.
Structura ARN mesager sau de informatie. Se numeste astfel intrucat , transporta mesajul, informatia continuta in ADN. Ca atare ARNm este deplasat din nucleu in citoplasma la locul sintezei proteice, unde se ataseaza de ribozomi alcatuind polisosomi activi. In deplasare in citoplasma ARNm este insotit de proteine care ii asigura stabilitatea. Formeaza cu aceste proteine complexe nucleoproteice numite informozomi. ARNm are o durata scurta de viata. Cu toate ca se distruge repede, elk este dintetizat mereu la nivelul cromozomilor in functie de necesitatile celulei.
Structura ARN ribosomal este monocatenara ca si ARNv. Exista mai multe feluri de ARN in functie de unitatile sau subunitatile ribozomale in care intra
Desi toate tipurile de ARNr sunt monocatenare, ele prezinta numeroase portiuni fals bicatenare, numite "agrafe de par". Aceste portiuni se realizeaza pe seama bazelor azotate complementare din segmentele vecine ale aceleiasi monocatene.
Bazati pe cunoasterea structurii ARNr si a proteinelor ribozomale precum si pe observatia ca la suprafata ribizomilor se afla o mare cantitate de ARNr, au propus un model de distributie spatiala a celor 2 tipuri de substante la nivelul ribozomului.
ARNr nu se poate prezenta ca o structura bicatenara complementara, ci numai portiuni fals bicatenare, realizate pe seama aceleasi catene.
Implicatiile ARNr in realizarea mecanismelor ereditare nu sunta inca pe deplin elucidate.
 Structura
ARN de transport (ARNt) sau solubil (ARNs) este
tot monocatenara, dar are o forma specifica de trifoi cu 4 foi si o structura
tridimensiunala ce aminteste structura proteinlor. Rolo ARNt este de a se cupla cu aminoacizii si
de a-I transporta la locul sintezei
proteice. In general o molecula de ARNt cuprinde un
numar mic de nucleotide, avand o greutate moleculara relativ mica.
Structura
ARN de transport (ARNt) sau solubil (ARNs) este
tot monocatenara, dar are o forma specifica de trifoi cu 4 foi si o structura
tridimensiunala ce aminteste structura proteinlor. Rolo ARNt este de a se cupla cu aminoacizii si
de a-I transporta la locul sintezei
proteice. In general o molecula de ARNt cuprinde un
numar mic de nucleotide, avand o greutate moleculara relativ mica.
Acizii ribonucleici de transport prezinta acelasi plan general de structura, determinate de felul succesiunii nucleotidelor sale
ARNt prezinta in structura monocatenara baze minore (inozina, pseudouiridina, dihidrouridina, baze de sulf - tiouridina), baze care nu prezinta o specificitate de complementaritate cu bazele obisnuite, precum si usoara neconcordanta cantitativa intre bazele obisnuite, complementare, determina structura specifica a ARNt.
Monocatena se infasoara secundare prin imperecherea bazelor azotate complementare din diverse zone, rezultand in final , 4 portiuni de pseudohelix, numite tulpini (tulpina acceptor, anticodon, D si T), separate de 4 bucle
DENATURAREA SI RENATURAREA ACIZILOR NUCLEICI; FORMAREA DE HIBRIZI MOLECULARI
DENATURAREA prin caldura - Daca ADN-ul este suficient de incalzit, portiunile sale dublu-catenare se desfac datorita ruperii legaturilor de H intre bazele azotate complementare. In acest caz ARN devine monocatenar pe intreaga sa lungime. Prin acest tratament poate fi denaturata si molecula dublu catenara a ADN-ului. La o racire treptata a solutiei bazele azotate isi pot reface legaturile de H si ADN-ul devin din nou bicatenar numin-du-se ADN renaturat. Si in cazul ARN-ului, la o racire treptata a solutiei pot reaparea portiuni dublu catenare.
Acest fenomen de denaturare-renaturare este utilizat pentru formarea de hibrizi moleculari, cat si pentru cercetarea secventionalitatii bazelor azotate, de-a lungul macromoleculelor de acizi nucleici.
Caracteristici fizico-chimice a ADN monocatenar nativ si/sau denaturat:
|
Carcateristici |
ADN nativ |
ADN denaturat |
|
Absorbtia raze UV |
Cantitate similara de raze cu lungimea de unda 2600 A |
+40% radiatii UV mai mult |
![]() profilul
de denaturare sau punctul de topire a ADN-ului
profilul
de denaturare sau punctul de topire a ADN-ului
Fenomenul de renaturare asigura formarea de hibrizi moleculari atat intre 2 ccatene de AND cat si intre o catena de ARN si una de ADN. Conditia este ca cele doua catene care hibrideaza, sa prezinte suficiente portiuni complementare. Astfel de experiente de hibridari moleculare au demonstrat ca ARN-ul unui organism este in totalitate complementar cu ADN-ul organismul respective. Posibilitatea de realizarea a hibrizilor moleculari ADN-ARN sau ADN-ADN intre specii, este cu atat mai mare cu cat speciile respective sunt mai apropiate din punct de vedere filogenetic.
Daca doua catene de ADN apartinand la specii diferite sunt complementare, inseamna ca prima catena este identica cu complementara celei de a doua. Pe aceasta baza se pot stabili deci portiunile de ADN de la specii diferite care porta aeeasi informatie ereditara. Portiunile renaturate pot fi decelate nu numai prin tehnica elctronico-microscopica, ci si prin tehnica radiugrafiei. In acest caz una din catenele utilizate in experiment, trebuie sa detina un izotop radioactive. In acest fel zonele unde s-a realizat renaturarea , vor imprima placa fotgrafica.
ORGANIZAREA ARN SAU ADN IN CROMOZOMUL VIRAL
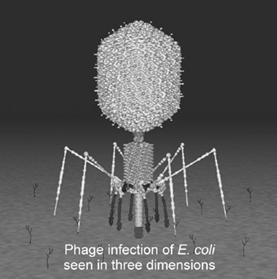 Materialul genetic al ARN-virusurilor prezinta diverse aspecte de
organizare si condensare. Astfel ARN-ul VMT este
monocatenar prezentand un aspect linear. Unele forme foarte mici de ARNv poseda
tot o macromolecula monocatenara de ARN, dar aceasta pe unele portiuni, devine
secundar fals dublu-catenara.acidul nucleic prezentand o forma spatiala cu mai
multe brate duble.
Materialul genetic al ARN-virusurilor prezinta diverse aspecte de
organizare si condensare. Astfel ARN-ul VMT este
monocatenar prezentand un aspect linear. Unele forme foarte mici de ARNv poseda
tot o macromolecula monocatenara de ARN, dar aceasta pe unele portiuni, devine
secundar fals dublu-catenara.acidul nucleic prezentand o forma spatiala cu mai
multe brate duble.
In cazul virusurilor ADN, materialul genetic se prezinta de asemenea, sub diverse aspecte. De exmplu molecula de ADN a virusului phiM13 este monocatenara liniara, iar a virusului phi X174 este tot monocatenara dar circulara.
Printre virusurile ce poseda ADN dublu-catenar se disting mai multe variante:
ORGANIZAREA ADN LA CELULELE PROCARIOTE
Ca si virusurile, organismele cu structura celulara prezinta diverse modalitati de impachetare a materialului genetic. La procariote, la E.coli , uriasa molecula circulara de ADN prezinta 12-80 de bucle, fiecare bucla prezentand numeroase superrasuciri, fiecare alcatuite din aproximativ 400 de perechi de nucleotide.
Bacteriile mai detin in afara cromozomului circular, material genetic sub forma de plasmide. Aceasta sunt structuri circulare de ADN cu capacitate de autoreproducere. Ele sunt structuri circulare de ADN cu capacitate de reproducere. Ele reprezinta aproximativ 1% din lungimea cromozomului bacterioan.
ORGANIZAREA ADN LA EUCARIOTE
Celulele eucariote detin materialul genetic nuclear sub forma de cromozomi. Cromozomii celulelor tinere (inainte de faza de autoreplicare a ADN-ului), detin o singura macromolecula de ADN bicatenar alcatuind cromozomii unicromatidici. Dupa faza S de replicare a ADN-ului, cromozomii devin bicromatidici posendand doua macromolecule de ADN bicatenar.
Macromolecule de ADN sunt strans plicaturate la nivelul cromozomilor datorita interactiunii cu proteinele histonice formand complexe nucleo-proteice. Aceste complexe se datoresc interactiunilor dintre sarcinile negative ale radicalilor fosforici ai ADN-ului sai sarcinile positive ale aminoacizilor din proteinele histonice de natura bazica. Plicatura fibrei, poate fi transversala, longitudinala sau combinata.
Picaturarea combinata determina superrasuciri asigurand condensarea materialului genetic in timpul diviziunii celulare, atat a cromozomilor cromatidici, cat si la cei unicromatidici. In urma cercetarilor s-a stability ca fibra de cromatina este alcatuita din nucleozomi dispusi cap la cap.
Nucleozomul este unitatea fundamentala de organizare a fibrei de cromatina. El are forma unui cilindru turtit. Fiind format din proteine histonice. Aceste proteine au greutati moleculare mici. La toate organismele eucariote ele se impart in 5 clase :
|
Denumirea |
Greutatea Moleculara |
Aminoacizi preponderenti |
Variatia compozitiei in aminoacizi la diferite specii |
|
H1 |
Lizina |
Varabila |
|
|
H2A |
Lizina |
Relativ stabila |
|
|
H2B |
Lizina |
Relativ stabila |
|
|
H3 |
argintina |
Foarte stabila |
|
|
H4 |
argintina |
Foarte stabila |
Miezul nucleozomului este format din 8 molecule histomice. Molecula de ADN infasoara de doua ori cilindrul histonic, apoi trece in urmatorul cilindru realizand din nou 2 infasurari.
PROBLEMA ORGANIZARII SUPRACROMOZOMIALE A MATERIALULUI GENETIC IN NUCLEUL CELULELOR EUCARIOTE
Daca fiecare cromozom unicromatidic are o singura molecula de ADN, iar unul bicromatidic are doua , ar insemna ca in nucleul eucariotelor exista atatea molecule de ADN cate cromatide sunt. O serie de cercetari vin sa infirme acest punct de vedere. Astfel, s-a remarcat ca 12 din cei 14 cromozomi de la Oenothera lamarkiana sunt legati intre ei formand un mare. Cercetarile electronomicroscopice au confirmat aceasta supozitie. Astfel de cercetari au permis sustinerea ipotezei ca celulele eucariote detin in nucleul lor o singura macromolecula de ADN, iar dupa faza de replicare a ADN-ului ar exista doua astfel de molecule.
CONCLUZII
Strucutra moleculara a tuturor tipurilor de ARN este monocatenara, unele tipuri prezentand si portiuni fals bicatenare, datorate bazelor azotate.
ADN-ul detine riboza ca zahar, iar bazele azotate sunt A, T, G, C. Structura sa este de obicei bicatenara, cele doua catene fiind antiparalele.
Natura informatiei genetice este data de secventa si numarul perechilor de baze azotate.
Aberts B, Bray
D, Lewis, Watson (1985) - Molecular Biology of the cell,
Nicolae Coman (2003) - Genetica Volumul 1, BCU, Cluj - Napoca
Bahr, G.F>
(1977) - Chromosomes and Cromatin Structure, Acad,
Benzer S (1955),
Fine Structure of a
Gene Region in Bacteriophage, Proc..Nat. Acad. Of Sciene
Sursa imaginilor www.google.com
Universitatea Babes Bolyai
Facultatea de Biologie si geologie
Anul II , biologie , grupa 3
Turony Alexandra Mihaela
|
Politica de confidentialitate |
| Copyright ©
2024 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |
