
aciziI ribonucleici
Structura si clasificarea acizilor ribonucleici
Acizii ribonucleici (ARN) sunt molecule poliribonucleotidice, rezultate din polimerizarea a patru tipuri de ribonucleotide, in care bazele azotate sunt A, U, C, G. Fiecare ribonucleotid contine o pentoza - riboza, o baza azotata si legaturi fosforice. In timpul sintezei ARN, incatenarea ribonucleotidelor se face cu ajutorul ARN-polimerazei.
Se disting doua categorii de ARN, celulare si virale. ARN-urile celulare nu codifica informatie genetica, dar sunt implicate in transcrierea si translatia mesajelor genetice; ARN-urile virale insa, reprezinta substratul ereditar, codificat al ribovirusurilor.
Moleculelor de ARN li se descriu:
structura primara, monocatenara, reprezentata de secventializarea nucleotidelor si stabilizata prin punti fosfodiesterice;
structura secundara, partial bicatenara in unele zone in care bazele A-U si C-G sunt legate complementar, intracatenar;
structura tertiara, rezultata prin asezarea intr-o configuratie tridimensionala a catenei ribonucleotidice.
ARN-urile celulare se clasifica in:
ARN mesager (ARN-m);
ARN-uri transportoare (ARN-t);
ARN ribosomal (ARN-r).
ARN mesager (ARN-m)
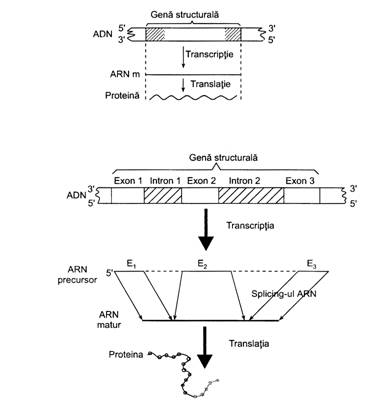
ARN-m este un poliribonucleotid monocatenar, sintetizat in nucleu dupa modelul unei catene a ADN-ului (in mod complementar). Prin sinteza lui conform codonilor din gena, preia (copiaza) informatia genetica, fiind in acelasi timp si purtator al mesajului genetic pana in citoplasma, la nivelul ribosomilor, in vederea sintezei de proteine.
Dat fiind ca sinteza ARN-m reprezinta de fapt mecanismul transcriptiei genice, structura ARN-m este heterogena, proprie, dictata de structura genei de pe care s-a sintetizat; lungimea moleculelor de ARN-m variaza de asemenea, in functie de marimea mesajului genic copiat (transcris) din ADN.
Sinteza ARN-m are loc in interfaza ciclului celular. Initial se sintetizeaza fragmente de ARN-m (sau preARN-m) care copiaza mesajul - secventializarea nucleotidelor - din exoni si introni. Apoi urmeza un proces de prelucrare post-transcriptionala, de matisare (asamblare), in care sunt excizate portiunile sintetizate de la nivelul intronilor, iar cele de pe exoni se sudeaza cap la cap, printr-o alegere multipla variabila (1,2,3,4,..sau 1,3,2,4,..s.a.m.d.). In felul acesta rezulta molecula de ARNm matur, detinatoare a unui anumit mesaj genetic.
Prin acoperirea moleculei de ARN-m de catre proteine, se asigura protectia acesteia fata de actiunea endonucleazelor.
ARN-m matur odata format, migreaza in citoplasma la nivelul ribosomilor, unde va avea loc procesul decodificarii mesajului genetic (intre codonii ARN-m si anticodonii ARN-t). Durata de viata a ARN-m este variabila, de 2-4 minute la procariote si 1-12 ore la eucariote, dupa care se va dezintegra. In situatii speciale, cum este cazul reticulocitelor la mamifere, ARN-m necesar pentru sinteza hemoglobinei este sintetizat anterior disparitiei nucleului, ramanand activ ulterior, cateva zile.
ARN-urile transportoare (ARN-t)
ARN-t prin functia pe care o indeplineste mai este numit si ARN adaptator sau acceptor de aminoacizi. El se afla in citoplasma intr-o proportie de aproximativ 15-18% din totalul ARN-ului celular; are o durata lunga de viata si buna stabilitate incat, o aceeasi molecula este folosita de repetate ori in translatie.
Genele pentru sinteza ARN-t sunt grupate in structura ADN-ului, in copii multiple, ocupand zonele telomerice si cele din constrictiile secundare ale cromosomilor acrocentrici.
In citoplasma celulelor s-au descris pana in prezent peste 40 de tipuri de ARN-t. Fiecare molecula de ARN-t este compusa dintr-o catena poliribonucleotidica, formata prin insiruirea a 70-95 ribonucleotide (aceasta reprezinta structura primara).
ARN-t prezinta si structura secundara, respectiv tertiara, catena fiind infasurata tridimensional in forma unei frunze de trifoi (cloverleaf). Din aceasta configuratie rezulta cateva regiuni si anume:
o regiune denumita acceptoare, formata intotdeauna din secventele CCA si care reprezinta locul (situs) de atasare si transport al unui aminoacid activat;
o regiune (bucla) formata din 7-12 nucleotide ce reprezinta situs-ul de recunoastere al ribosomilor si de legare a ARN-t de ribosomi;
o regiune (bucla) variabila, denumita 'ciot', a carei functie nu este cunoscuta;
o regiune (bucla) formata din 7 nucleotide, din care cele 3 baze azotate centrale formeaza un 'anticodon'. Structura secventiala a anticodonilor variaza de la un ARN-t la altul;
o regiune (bucla) formata din 6-12 nucleotide, reprezentand locul de recunoastere si legare a enzimei amino-acil-sintetaza de ARN-t. In citoplasma celulelor exista peste 20 de tipuri de astfel de enzime, ele avand rol activator in procesul de legare si transport al aminoacidului, deci in procesul translatiei.
ARN ribosomal (ARN-r)
ARN-r reprezinta aproximativ 80% din totalul ARN-ului celular si 2% din componenta ribosomilor. Sinteza sa se face pe baza mesajelor codificate de genele ribosomale, situate in constrictiile secundare si in telomerele cromosomilor. Structura primara este reprezentata de secventializari in numar mare de ribonucleotide. In molecula de ARN-r alterneaza zone monocatenare cu zone bicatenare (structura secundara), molecula prezentand si o configuratie tridimensionala caracteristica (structura tertiara).
ARN-r este intotdeauna legat de proteine ribosomale impreuna cu care formeaza scheletul structural al ribosomilor.
Expresia fundamentala a genelor (transferul informatiei genetice)
Dogma centrala a geneticii moleculare
In 1958, F. Crick a emis ipoteza transmiterii unidirectionale a mesajului informational genetic de la ADN la proteine, prin intermediul ARN-m, mentionand ca este imposibil transferul de informatie de la ADN direct la proteine si in sens invers. Aceasta afirmatie a devenit unanim acceptata, fiind supranumita 'Dogma centrala a geneticii moleculare' si notata sintetic:
 Transcriptie Translatie
Transcriptie Translatie
ADN ARN-m proteina
Replicare
semiconservativa
In 1965, Watson aduce 'dogmei centrale' unele completari si precizari referitoare la imposibilitatea transmiterii informatiei genetice de la ARN la ADN, deoarece ARN nu ar putea functiona ca model (matrita) (fig.14a si 14b).
In aceeasi perioada de timp insa, Temin studiind mecanismul de oncogeneza virala prin retrovirusuri (virusuri cu ARN), avanseaza o noua ipoteza de transcriere a informatiei genetice de la ARN-ul viral la ADN. Ipoteza facea referiri directe la un posibil mecanism de formare a unui ADN monocatenar, complementar cu ARN-ul viral si prin care s-ar permite integrarea retrovirusurilor in ADN-ul gazdei.
Confirmarea acestei ipoteze a fost facuta in 1970 de catre Temin si Baltimore (pentru care au fost rasplatiti cu premiul Nobel), care au demonstrat existenta transcrierii inverse a informatiei genetice, de la ARN la ADN in cazul virusurilor cu genom constituit din ARN. In acelasi timp s-a descoperit si existenta in genomul retrovirusurilor si a celulelor canceroase a enzimei revers-transcriptaza (sau transcriptaza inversa), care joaca rol important in transferul de informatie de la ARN la ADN si in polimerizarea noii catene de ADN.
Transferul informatiei de la ADN ARN proteine
Descoperirea transcrierii inverse a dus inerent la revizuirea si reformularea dogmei centrale. Astfel, in 1970, F. Crick concluzioneaza ca transcrierea inversa nu infirma dogma centrala, dar ca aceasta din urma necesita o reformulare, respectiv o noua viziune asupra posibilitatilor de transfer informational, sumarizata astfel:
transfer general, prezent in toate celulele cu genom ADN, care se face: ADN ARN proteine;
transfer special, prezent numai in anumite circumstante, de la
ARN ADN ARN-m proteine;
transfer (mai putin cunoscut) de la proteina la ADN, prin produsii de sinteza ai unor gene structurale, care interactioneaza specific cu ADN-ul in reglajele genice.
Transcriptia mesajului genetic (Biosinteza ARN-m)
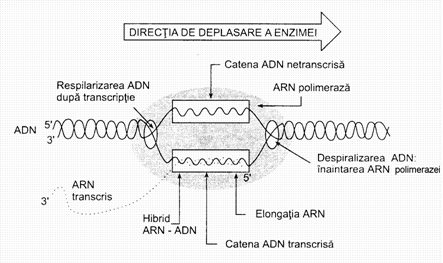
Transcriptia este un proces complex, de sinteza a unei molecule de ARN-m folosindu-se drept matrita una dintre cele doua catene ale ADN-ului, intotdeauna aceeasi, datorita antiparalelismului catenar din structura ADN-ului.
Sinteza se face conform complementaritatii bazelor azotate ale ADN-ului cu cele din ARN-m, incat prin sinteza unei molecule de ARNm se copiaza (transcrie) mesajul genetic codificat in portiunea de ADN folosita drept matrita. Intotdeauna este copiata o singura catena a ADN-ului si anume cea cu legaturile fosforice orientate in sens 3' - 5' .
Modul de desfasurare al transcriptiei
Biosinteza ARN-m (transcriptia) se desfasoara in prezenta enzimei ARN-polimeraza (transcriptaza). Aceasta enzima prezinta cateva subunitati, necesare functiei sale, dintre care amintim:
o subunitate care asigura recunoasterea zonei promotorului (zona de unde incepe citirea genei);
o subunitate care asigura buna desfasurare a sintezei de ARN-m.
Transcriptaza (ARN-polimeraza) este capabila sa identifice catena care urmeaza a fi transcrisa, de asemenea capatul de unde incepe citirea si capatul terminal al genei prin legarea sa reversibila de proteine specifice. In absenta acestora, initierea citirii genei este gresita, la fel asamblarea si lungimea catenei de ARN-m. Este astfel bine cunoscut ca aceste proteine servesc la recunoasterea codonilor 'stop', in vederea sistarii sintezei ARN-m si eliberarii acestuia de pe matrita.
In mecanismul transcrierii au loc:
faza de pre-initiere sau de "cautare" a regiunii promotor de catre ARN-polimeraza (transcriptaza);
faza de initiere, cand in prezenta unei enzime se desfac legaturile de hidrogen ale ADN, iar de pe una din catene se incepe citirea - asezarea ribonucleotidelor- conform complementaritatii;
faza de elongare, in care transcriptaza aluneca de-a lungul monocatenei de ADN, polimerizand antiparalel ribonucleotidele in sens 5' - 3'. Pe masura ce transcriptia avanseaza, ADN isi reia progresiv structura dublu catenara. Procesul transcriptiei se desfasoara cu o viteza de 60 de nucleotide pe secunda, timpul variind in functie de tipul genei si de diversi factori ce actioneaza in acel moment;
faza de terminare, care corespunde cu recunoasterea codonilor stop si deci, stoparea transcriptiei, cu desprinderea ARN-m.
Transcriptia este un proces selectiv, copiindu-se - intr-un moment precis- mesajul de pe o singura gena din genomul celular.
Dupa incheierea transcriptiei, ARN-m va suferi un proces de indepartare a fragmentelor sintetizate de pe introni si apoi de legare (splicing) a fragmentelor sintetizate de pe exoni, incat in final se formeaza molecula de ARN-m matur. In situatia in care se produce o "greseala" in procesul de excizie al zonelor transcrise de pe introni, se va sintetiza o proteina cu efecte patologice. Un astfel de exemplu, este miodistrofina intalnita in miodistrofia Duchenne.
Dupa cum s-a mentionat intr-un capitol anterior, la procariote genele au structura liniara, incat transcriptia unui operon se face indeobste coliniar (un operon este un grup de gene structurale coordonate in comun de o gena de control).
La eucariote insa, genele au structura mozaicata (fiind formate din secvente traductibile -translabile- denumite exoni si netranslabile, denumite introni); prima molecula de ARN-m (cu transcriptul primar) urmeaza a inlatura fragmentele sale sintetizate de pe introni, iar cele de pe exoni vor fi ulterior legate, asamblate. Asamblarea poate fi cu alegere multipla existand mai multe posibilitati ca de pe aceeasi gena sa fie transferata informatia ereditara pentru sinteza mai multor tipuri de proteine, proces numit alternanta de matisare.
In urma asamblarii fragmentelor ia nastere ARN-m matur care se asociaza cu proteine, migreaza in citoplasma si se leaga de ribosomi (intre unitatea mare si cea mica), in vederea procesului de translatie.
Translatia mesajului genetic in biosinteza proteinelor
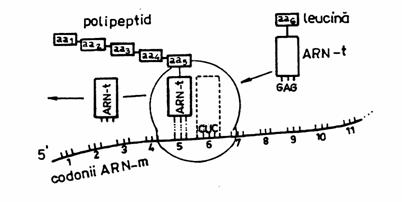
Translatia si sinteza proteinelor au loc in ribosomi. Acestia sunt pe drept cuvant uzinele de biosinteza proteica a celulelor.
Ribosomii se leaga reversibil de ARN-m si ARN-t incarcat cu aminoacid, astfel incat sunt utilizati de repetate ori in traducere. Ei joaca rolul de a orienta molecula de ARN-m in vederea bunei translari intre codon - anticodon.
Translatia este reprezentata de traducerea codonilor din ARN-m (respectiv codonii transcrisi din ADN) de catre anticodonii ARN-t, incat informatia codificata prin alfabetul codului genetic este convertita intr-un alfabet al celor 20 de aminoacizi.
Pentru efectuarea traducerii mesajului din ARN-m in vederea sintezei proteice este necesara prezenta de: aminoacizi liberi, ARN-m matur, ribosomi, ARN-t, enzime amino-acil-sintetaze (cu rol in formarea complexului adaptator ARN-t-aminoacid), mitocondrii ca surse de energie etc.
Mecanismul translatiei
Mecanismul translatiei se desfasoara in urmatoarele faze:
faza de activare a aminoacizilor;
faza de initiere;
faza de elongare a polipeptidului;
faza de terminare a traducerii mesajului.
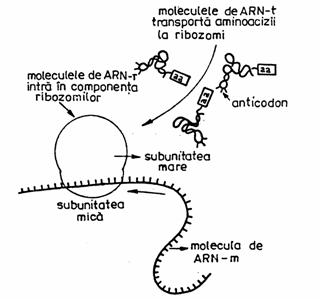
Rolul celor trei tipuri de ARN in procesul de translatie.
Activarea aminoacizilor (AA) consta in legarea fiecaruia de o molecula de ARN-t si de o enzima amino-acil-sintetaza, constituind impreuna complexul adaptor, activ. In celule exista cel putin 20 de tipuri diferite de amino-acil-sintetaza, cate una pentru fiecare din cei 20 de AA.
Faza de initiere implica cateva procese, in care rol principal il are codonul initiator (codonul care specifica metionina) (vezi in continuare 'Codul genetic').
In faza de elongare, decodificarea intre codon-anticodon continua de-a lungul ARN-m matur, succesiv. In paralel cu asezarea AA incepe si polimerizarea lor, prin formarea de legaturi peptidice intre AA.
Faza de terminare se caracterizeaza prin sistarea translatiei, la aparitia unuia din cei trei codoni 'stop'.
Codul genetic
Incepand cu deceniul al V-lea al secolului XX, cercetarile de genetica moleculara au permis stabilirea existentei codului genetic pentru toate vietuitoarele si treptat, cunoasterea in intregime a mecanismelor acestuia. Astfel, F.Crick si colaboratorii (1961), pe baza datelor acumulate in genetica moleculara, descriu caracteristicile codului genetic, stabilind ca trei nucleotide alaturate constituie un codon (triplet) care reprezinta o unitate a codului genetic, de specificare a cate unui aminoacid pentru sinteza proteinelor. Practic, in 1965, studiile mai multor grupuri de cercetatori au permis cunoasterea in intregime a dictionarului codului genetic din structura genelor.
Termenul de cod genetic inglobeaza sistemul de corespondenta intre codoni (fiecare codon cuprinde 3 nucleotide adiacente din ADN) si aminoacizii din structura proteinelor. Cu alte cuvinte, codul genetic este forma de depozitare si conservare a informatiei genetice, un adevarat dictionar bilingv prin care literele - respectiv cele patru tipuri de nucleotide: A, T, C, G - sub forma de cuvinte (codoni), servesc la specificarea (asezarea) aminoacizilor in proteine. Fiecare codon este strict specific.
Codonii (tripletele) reprezinta cuvintele codului genetic; ei sunt formati din trei nucleotide adiacente din ADN, respectiv din ARN-m (prin transcriptie); fiecare codon va corespunde unui aminoacid din proteina.
Primele experimente efectuate pentru descifrarea codului genetic au constat in sinteza unui ARN-mesager (ARN-m) poliuracilic, ale carui secvente de nucleotide contineau ca baza azotata doar uracil-ul (U). Acest ARN-m sintetizat, adaugat la un sistem activ de ribosomi, in prezenta tuturor tipurilor de aminoacizi, a ARN-urilor transportoare (ARN-t) si a enzimelor activatoare, a dus la sinteza unui lant polipeptidic compus exclusiv din fenilalanina (polifenilalanina); pe baza acestor experimente s-a conchis ca tripletul pentru fenilalanina este UUU.
Descifrarea integrala a codului genetic a fost posibila din 1965, pe baza experimentelor lui Nierenberg, Ochoa si Khorana, ei avand meritul de a fi stabilit semnificatia celor 64 de codoni (43=64). Din acestia, 61 sunt codoni cu 'sens', specificand cei 20 de aminoacizi, iar trei codoni, denumiti 'fara sens' au rol de codoni 'stop' sau 'terminatori' ai sintezei proteice.
|
U |
C |
A |
G | ||||||
|
U |
UUU |
phe |
UCU |
ser |
UAU |
tyr |
UGU |
cys |
U |
|
UUC |
phe |
UCC |
ser |
UAC |
tyr |
UGC |
cys |
C |
|
|
UUA |
leu |
UCA |
ser |
UAA |
stop |
UGA |
stop |
A |
|
|
UUG |
leu |
UCG |
ser |
UAG |
stop |
UGG |
try |
G |
|
|
C |
CUU |
leu |
CCU |
pro |
CAU |
his |
CGU |
arg |
U |
|
CUC |
leu |
CCC |
pro |
CAC |
his |
CGC |
arg |
C |
|
|
CUA |
leu |
CCA |
pro |
CAA |
gln |
CGA |
arg |
A |
|
|
CUG |
leu |
CCG |
pro |
CAG |
gln |
CGG |
arg |
G |
|
|
A |
AUU |
ile |
ACU |
thr |
AAU |
asn |
AGU |
ser |
U |
|
AUC |
ile |
ACC |
thr |
AAC |
asn |
AGC |
ser |
C |
|
|
AUA |
ile |
ACA |
thr |
AAA |
lys |
AGA |
arg |
A |
|
|
AUG + |
met |
ACG |
thr |
AAG |
lys |
AGG |
arg |
G |
|
|
G |
GUU |
val |
GCU |
ala |
GAU |
asp |
GGU |
gly |
U |
|
GUC |
val |
GCC |
ala |
GAC |
asp |
GGC |
gly |
C |
|
|
GUA |
val |
GCA |
ala |
GAA |
glu |
GGA |
gly |
A |
|
|
GUG |
val |
GCG |
ala |
GAG |
glu |
GGG |
gly |
G |
|
Codul genetic: combinatii posibile in triplete si semnificatia codonilor
Deoarece exista 64 de tipuri de codoni pentru specificarea celor 20 de aminoacizi rezulta ca un aminoacid poate fi specificat de mai multi codoni (denumiti si codoni sinonimi). Aceasta proprietate a codului genetic este cunoscuta sub numele de cod genetic degenerat. Pe baza acestei proprietati, eventualele mici mutatii (in structura codonilor) pot sa nu fie receptate, neantrenand modificari ale structurii proteinei respective.
Leucina si arginina pot fi specificate fiecare de 6 codoni sinonimi; alti aminoacizi pot fi specificati de 2-3 codoni. Numai metionina si triptofanul au fiecare un singur codon specific.
Initierea translatiei (a citirii ARN-m) este facuta de catre codonul corespunzator metioninei (codonul AUG). Se deduce deci ca primul aminoacid in lantul polipeptidic este metionina. La incheierea sintezei, dupa caz, metionina se poate desprinde din lantul polipeptidic.
Trei dintre codonii codului genetic, asa numitii 'codoni stop' (UGA, UAA, UAG) au rolul de oprire a translatiei de pe ARN-m.
Codul genetic prezinta urmatoarele proprietati:
este tripletic; astfel, un codon semnifica un aminoacid; din cei 64 de codoni, 61 sunt codoni cu sens, specificatori de AA, iar 3 codoni coordoneaza oprirea citirii;
este degenerat: o parte dintre codonii cu sens sunt sinonimi, specificand un acelasi tip de aminoacid;
este nesuprapus (nesuperpozabil): aceasta caracteristica se refera la faptul ca doi codoni succesivi nu au in mod normal nici un nucleotid comun;
este inscris fara 'virgule', codonii nefiind separati intre ei (nu exista structuri nucleotidice care sa-i separe);
este lipsit de ambiguitate, ca atare un codon dat specifica un singur aminoacid, intotdeauna acelasi;
este inscris si citit liniar; o eventuala pierdere de nucleotide va duce la citirea eronata de la locul mutatiei, determinand sinteza de proteina patologica, in 'frame-shift';
dintr-un codon, primele doua nucleotide joaca rol mai important, cea de-a treia putand fi schimbata (oscilanta), fara a perturba sinteza si structura proteinei;
este universal; aceasta proprietate se refera la faptul ca pentru toate speciile, aminoacizii sunt specificati de aceleasi tipuri de codoni.
Proprietatea de cod universal atesta originea comuna a vietuitoarelor pe scara animala. Cu toate acestea, in cadrul diferitelor specii, se remarca totusi o utilizare preferentiala a anumitor codoni, in functie de necesarul de proteine specifice fiecarei specii.
Cercetari privind genomurile virale au descoperit ca la unele virusuri (printre care si virusul HIV care determina SIDA) exista acoperiri -suprapuneri- de gene, acestea reprezentand probabil posibilitatea de folosire cu maxima eficienta a unui genom amplasat intr-un spatiu restrans. Cu alte cuvinte, acoperirea de gene se refera la faptul ca o gena detine capacitatea de codificare multivalenta, situatie care este intalnita la virusuri. Unele suprapuneri partiale se gasesc si la genele mitocondriale.
Baza genetica a sintezei si structurii proteinelor.
Relatia gena-proteina
La organismele eucariote proteinele sunt sintetizate la nivelul ribosomilor, pe baza programelor informationale vehiculate de la nivelul ADN-ului, prin intermediul ARN-mesager (ARNm).
Sinteza proteica (modul de secventializare a aminoacizilor - pozitia - fiecaruia) este dictata de informatia codificata in ADN; structura fiecarei proteine, in special structura primara, confera o anume functionalitate proteinei respective.
Multiple studii de genetica moleculara au demonstrat ca functionalitatea, specificitatea, fiecarei proteine este conferita de catre fiecare aminoacid, mai exact de pozitia ocupata de acesta in lantul polipeptidic, guvernata de mesajul genetic.
Orice modificare a secventializarii unuia sau mai multor aminoacizi atrage dupa sine modificari in structura proteinelor, insotite de modificari ale functionalitatii, destul de grav exprimate fenotipic.
In unele situatii in care, conform proprietatii de cod degenerat, in mutatia survenita codonii sunt sinonimi codonilor initiali, se vor secventializa aceleasi tipuri de aminoacizi, astfel incat proteina nu va suferi modificari; este cazul mutatiilor silentioase.
Este bine cunoscut ca toate caracterele fenotipice ale fiecarui organism sunt determinate de tipul de proteine, deci de programul genetic; proteinele sunt suportul biochimic al caracterelor fenotipice.
Proteinelor li se descriu: structura primara, secundara, tertiara iar pentru unele proteine chiar structura quaternara.
Structura primara este reprezentata de secventializarea aminoacizilor (pozitiile aminoacizilor in lantul polipeptidic). Aceasta structura dictata genetic prin transferul de mesaj genetic codificat pana la ribozomi, confera o inalta specificitate fiecarei proteine. Orice greseala in asamblarea -asezarea- aminoacizilor va perturba, desigur, structura, si, in acelasi timp, functia proteinei.
Specificitatea proteinelor mai este determinata si de configuratia lantului polipeptidic in spatiu (structura secundara), de legaturile chimice ce se stabilesc intre aminoacizi in cadrul aranjamentului tridimensional (structura tertiara) si, in fine, de structura quaternara, aceasta fiind caracteristica numai unor proteine (asocierea catorva lanturi polipeptidice, ca de exemplu Hb).
Efectele mutatiilor asupra structurii si functiei proteinelor
Secventializarea aminoacizilor fiind determinata de codonii din ADN, teoretic orice mutatie intervenita in gena ar putea avea drept consecinta:
modificarea structurii proteinei;
reducerea sau chiar abolirea sintezei proteice;
modificarea functionalitatii proteinei in sensul scaderii acesteia;
aparitia de functionalitati noi ale unei proteine modificata structural;
Efectele mutatiilor asupra structurii si functiei proteinelor pot determina, deci:
schimbarea (substitutia) unui aminoacid (de exemplu in hemoglobinopatii);
absenta unui aminoacid (ex. fibroza chistica - boala monogenica cu tulburari de transport membranal);
asezarea gresita a mai multor aminoacizi, datorita deletiei unui nucleotid (citire in frame-shift);
elongarea sau scurtarea unui polipeptid prin mutatii non-sens (mutatia codonilor stop) exemplu: hemoglobinopatia Constant Spring, talasemii etc;
scaderea functionalitatii proteinei si a capacitatii de sinteza a acesteia, consecutiv slabei functionalitati a intronilor, ceea ce duce la tulburari de asamblare a ARNm (exemplu: hemofilia A, neurofibromatoza etc);
abolirea sintezei proteinei in cazul mutatiei in promotor, prin abolirea sintezei ARNm. In acest caz gena este "silentioasa";
scaderea capacitatii de sinteza a proteinei sau chiar abolirea sintezei datorita unor multiple repetari secventiale (repetari nucleotidice); acestea reduc sau anihileaza sinteza de ARNm (exemplu: sindromul cu X-fragil, boala Huntington, miodistrofii etc.).
In contextul in care, conform proprietatii de cod genetic degenerativ, desi codonii au suferit mutatie, ei vor specifica aceleasi tipuri de aminoacizi (codonii cu mutatii sunt sinonimi cu precedentii) si mutatia nu se va exprima fenotipic. In acest caz se vorbeste de mutatie surda.
Relatia gena-proteina
Relatia directa dintre gena si proteina este evidenta in toate bolile moleculare, acestea fiind frecvente in patologia umana. Un exemplu edificator il constituie hemoglobinopatiile umane. In patologia umana sunt cunoscute peste 160 de hemoglobinopatii, ca rezultat al mutatiei genelor care specifica lanturile globinice.
In hemoglobinopatia S (anemia sicklica) mutatia punctiforma a genei beta in pozitia 6 a lantului beta-globinic produce substitutia acidului glutamic cu valina. Aceasta substitutie schimba profund functia Hb si chiar morfologia hematiilor care o contin, acestea capatand contur zdrentuit si tendinta mare de hemoliza. Consecutiv, se instaleaza anemie severa si alte tulburari clinice. Boala este raspandita pe toate continentele, avand insa o frecventa mai mare in Africa si zona mediteraneeana.
Hb-patia S in forma homozigota prezinta gravitate mai mare, fiind de obicei letala in primii ani de viata. In formele heterozigote, electroforetic se deceleaza 50% Hb A1 normala si 50% Hb S; aceasta reliefeaza ca alelele se afla in raport de codominanta.
In hemoglobinopatia C, in aceeasi pozitie a lantului beta, acidul glutamic este inlocuit cu lizina, consecutiv unei mutatii punctiforme in gena beta.
In hemoglobinopatia Constant Spring (Hb CS), lantul globinic contine zeci de insertii de aminoacizi in plus, datorita unei deletii a codonului stop, ceea ce duce la elongarea catenei a sau b
In hemoglobinopatia M, mutatia punctiforma a genei beta a lantului globinic produce substitutia tirozinei cu histidina.
Un alt grup de hemoglobinopatii este reprezentat de talasemiile a (foarte rare, fiind letale) sau b. Beta talasemiile pot fi determinate prin: substitutie de aminoacizi, deletii, mutatie non sens, defecte de asamblare a ARNm, citirea in frame-shift. Modificarile structurale ale lanturilor sunt dependente de tipul acestor mutatii genice si ele antreneaza tulburari grave ale functiei Hb respective.
Toate aspectele prezentate anterior subliniaza existenta corespondentei totale intre informatia codificata in gene si structura, respectiv functia proteinelor, ceea ce determina tocmai inalta specificitate de specie si de organism a proteinelor. Fiecare individ este un unicat in ceea ce priveste specificitatea proteinelor sale, singurele exceptii fiind gemenii monozigoti, ei avand proteine identice.
|
Politica de confidentialitate |
| Copyright ©
2024 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |