
BAZELE ANATOMO-FUNCTIONALE ALE SISTEMULUI NERVOS CENTRAL
Sistemul nervos are un rol extrem de important in structura oricarei forme de viata; acesta controleaza in totalitate orice miscare fara nici o exceptie. Sistemul nervos este cea mai complexa si cea mai importanta retea de control si de distribuire a informatiilor.
Structural sistemul nervos este constituit din 2 parți:
SNC (sistemul nervos central) care cuprinde encefalul si maduva spinarii, protejate de cutia craniana respectiv coloana vertebrala.
SNP (sistemul nervos periferic) care reprezinta restul de sistem nervos ce cuprinde ganglionii nervoși si nervii din tesuturi si organe. SNP nu este protejat de oase sau de o bariera pentru sange (fata de creier care este protejat de bariera hematoencefalica) si poate fi usor lezat mecanic sau chimic.
La randul lui sistemul nervos periferic este alcatuit din doua componente (Figura 1):
Sistemul nervos somatic caruia i se atribuie controlul voluntar al miscarilor prin actiunea muschilor scheletici cat si receptia stitmulilor externi (stimuli mecanici, stimuli termici etc). Sistemul nervos somatic este format din fibre aferente care duc informatiile de la surse externe catre SNC si fibre eferente care conduc impulsurile nervoase de la nivelul SNC catre mușchi.
Sistemul nervos autonom sau vegetativ: coordoneaza actiunile care nu se afla sub controlul constientului. Controleaza functii vitale precum respiratia, ritmul cardiac, dilatarea si constrictia pupilelor, respiratia, digestia etc. Acesta se imparte in sistem nervos simpatic si sistem nervos parasimpatic.
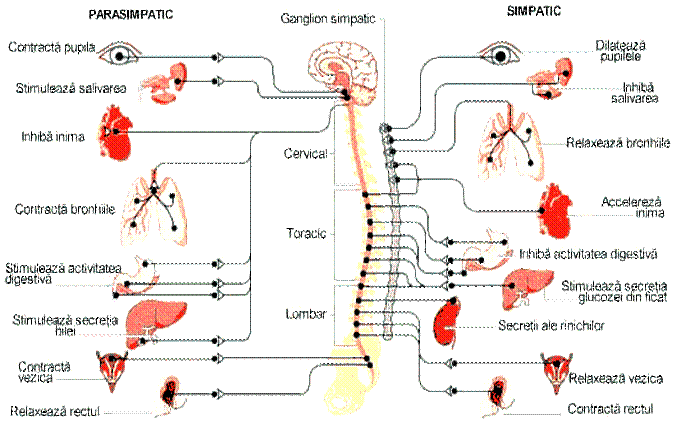
Figura 1
I.1. Encefalul
Encefalul (Figura 2) este situat in cutia craniana si in alcatuirea lui intra trunchiul cerebral, cerebelul, diencefalul si cele doua emisfere cerebrale, foarte dezvoltate la om, unde acopera aproape in intregime celelalte parti constitutive ale encefalului
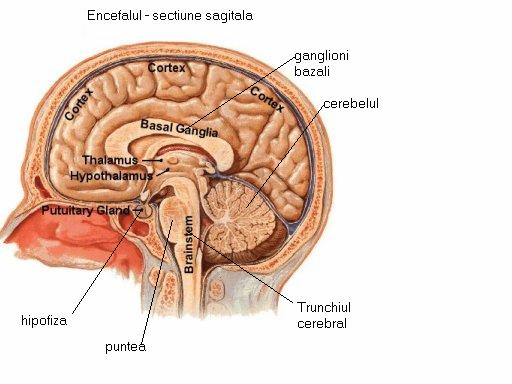 Figura 2
Figura 2
I.1.1. Trunchiul cerebral
Trunchiul cerebral continua maduva spinarii si este format din bulb, puntea lui Varolio si pedunculii cerebrali (mezencefal).
Trunchiul cerebral are aspectul unei coloane longitudinale cu doua fete una antero-laterala si una posterioara.
Este constituit din formatiuni cenusii si albe, omoloage celor medulare sau proprii fiecarui etaj. Substanta cenusie este situata la interior si fragmentata in numerosi nuclei, separati prin substanta alba care apare si la periferie.
La nivelul trunchiului cerebral se gasesc mai multi nuclei caracteristici fiecarui etaj al trunchiului (Figura 3) si avand functii specifice:
Nucleii bulbari:
nucleii motori: contin neuroni de origine ai fibrelor motorii ale nervilor cranieni, glosofaringian(IX), vag (X), accesor (XI), hipoglos (XII).
nuclei senzitivi: nuclei ce contin deutoneuronii cu care fac sinapsa fibrele senzitive ale nervilor cranieni glosofaringian (IX), vag (X), trigement (V), facial (VII) si nucleii vestibulari (VIII).
nuclei vegetativi: nucleul salivator inferior si nucleul dorsal al vagului care, impreuna cu cei din substanta reticulara sunt centrii unor reflexe viscerale controlate prin nervii glosofaringieni si vagi.
nuclei proprii: raspanditi in substanta reticulata sub sediul centrilor respiratorii, vasomotori si cardioinhibitori.
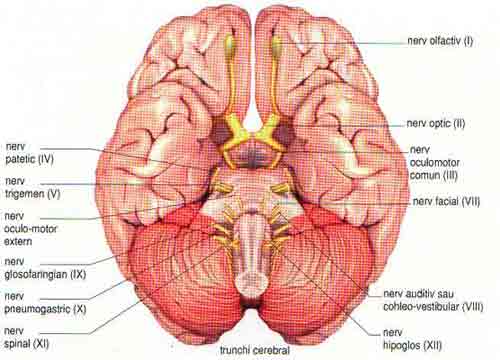 Figura 3
Figura 3
Nucleii pontini:
nuclei motori: ai nervilor cranieni trigemen (V), abducens (VI) si facial (VII).
nuclei senzitivi: ai nervilor cranieni VIII si nucleul principal senzitiv al nervului trigemen (V) .
nuclei vegetativi: salivator superior si lacrimal .
nuclei proprii: nucleii pontini si nucleii formatiunii reticulate.
Nucleii mezencefalici :
nuclei motori: ai nervilor cranieni oculomotor (III), trohlear (IV).
nuclei senzitiv: al tractului mezencefalic al nervului V.
nuclei vegetativi: accesori ai oculomotorului.
nuclei proprii: reprezentati prin substanta neagra care coordoneaza motilitatea asociata, dar mai ales intervin, impreuna cu scoarta cerebrala si hipotalamusul, in mecanismul somn-veghe.
nucleul roșu: are rol inhibitor asupra tonusului muscular.
Trunchiul cerebral este primul component al encefalului. Functiile sale sunt numeroase si de importanta vitala:
Funcția reflexa a trunchiului cerebral
Se realizeaza prin centrii din substanta cenusie, la acest nivel inchizandu-se o serie de reflexe de importanta vitala:
reflexele secretorii si motorii digestive.
reflexele respiratorii si adaptive cardiovasculare.
reflexul de glutitie.
reflexele salivare.
reflexele masticator, de clipire si mimica.
reflexele pupilar fotomotor si de acomodarea vederii la distanta.
reflexele statice si statokinetice, legate de mentinerea pozitiei corpului prin caile motoare extrapiramidale cu origine la acest nivel.
Functia de conducere a trunchiului cerebral
Se realizeaza prin substanta alba, alcatuita din fibre specifice ascendente, precum si din fibre descendente provenite din centrii nervosi superiori sau cu origine in trunchiul cerebral.
I.1.2. Cerebelul
Cea mai mare structura a creierului posterior este cerebelul. Aceasta zona are, in principal, activitati motorii. Trimite impulsuri care produc miscarile inconstiente ale muschilor, astfel postura si echilibrul sunt mentinute si actioneaza in perfect acord cu ariile motorii ale emisferelor cerebrale pentru coordonarea miscarilor corpului. Cerebelul, situat in etajul inferior al cutiei craniene, in spatele trunchiului cerebral, este alcatuit din doua emisfere cerebeloase (lobul anterior-paleocerebel si posterior-neocerebel), conectate printr-o parte mediana denumita vermis, unita cu o formatiune alungita, constituind arhicerebelul.
Arhicerebelul reprezinta creierul vestibular, fiind considerat ca centru al echilibrului gravitational prin participarea sa la coordonarea reflexelor de redresare labirintice si statokinetice. Astfel de implicatii functionale explica tulburarile de statica si mersul ebrios in cazul unor leziuni cerebeloase.
Paleocerebelul are un rol deosebit de important prin implicatia sa in reglarea tonusului muscular, a contractiei musculare de echilibru si de compensare oportuna a statiunii bipede sau a altor atitutidini posturale. Rolurile motorii ale paleocerebelului justifica tulburarile de tonus muscular precum: hipertofia, disparitia reflexelor de sustinere si redresare, identificate in leziunile cerebeloase si cunoscute sub numele de ataxie cerebeloasa'.
Neocerebelul considerat ca fiind dispozitivul regulator si coordinator al miscarilor in general, si al celor la nivelul extremitatilor, fiind responsabil de sicronizarea gesturilor si indirect a motricitatii voluntare. Aceste functii explica simptomatologia particulara a ataxiei cerebeloase, ca hipermetrie, tremuratura intentionala, opusa celei din parkinsonism, care este atenuata prin miscare.
I.1.3. Diencefalul
Diencefalul considerat creierul intermediar este localizat in prelungirea trunchiului cerebral, deasupra maxilarului, sub emisferele cerebrale. Este compus din talamoencefal si hipotalamus. Talamoencefalul cuprinde: talamusul, metatalamus, epitalamus, subtalamus.
Talamusul e compus din 2 formatiuni ovale de substanta cenusie dispuse lateral fata de ventriculul III. Talamusul reprezinta formatiunea cenusie diencefalica cea mai voluminoasa si cuprinde mai multi nuclei asociati cu trunchiul cerebral, cerebelul si scoarta cerebrala.
Talamusul are un rol hotarator in perceptia senzoriala, fiind ajutat de reglarea fina corticala.
Metatalmusul este alcatuit din corpii geniculati laterali si mediali; aici se afla centrul vizual primar. La acest nivel se termina tracturile optice care pleaca de la retina, iar eferentele lor merg in scoarta vizuala din lobul occipital. Corpii geniculati mediali sunt legati de caile auditive, de la ei pornesc radiatii acustice, care ajung la ariile auditive din lobul temporal. In concluzie metatalamusul are un rol semnficativ in ceea ce priveste transmiterea sensibilitatii vizuale si auditive.
Epitalamusul este alcatuit din glanda epifiza si un nucleu care influenteaza refexele olfactivo somatice (controleaza miscarile capului si mirosul).
Subtalamusul este constituit din fascicule de fibre si nuclei cenusii, are conexiuni cu toate celelalte formatiuni diencefalice, cu nucleul rosu si cu substanta neagra. Are rol in integrarea centrilor de control motor.
Hipotalamusul se afla la baza creierului, sub cele doua emisfere cerebrale. El este situat imediat sub o alta structura importanta din creierul anterior, talamusul, care functioneaza ca un releu telefonic intre maduva spinarii si emisferele cerebrale. Hipotalamusul este, in fapt, o colectie de centri nervosi specializati, care sunt conectati cu alte zone importante din creier si cu glanda hipofiza. Este regiunea creierului implicata in controlul unor functii vitale, cum ar fi mancatul, dormitul si termoreglarea. Este strans legat de sistemul hormonal endocrin.
Hipotalamusul are cai nervoase care il conecteaza cu sistemul limbic, care este strans legat de centrul olfactiv din creier. Aceasta portiune a creierului are, de asemenea, conexiuni cu arii ce controleaza alte simturi, comportamentul si organizarea memoriei.
I.1.4. Emisferele cerebrale
Sunt structuri nervoase, cunoscute si sub numele de creierul mare sau cerebrum. Acestea sunt mai dezvoltate la om decat la orice alt animal si sunt esentiale pentru gandire, memorie constienta si procesele mentale.
Reprezinta cea mai voluminoasa parte a encefalului Emisferele sunt unite la baza printr-un fascicul gros de fibre nervoase, denumite corp calos. Desi fiecare reprezinta imaginea in oglinda a celeilalte, ele au functii complet diferite si conlucreaza prin intermediul corpului calos
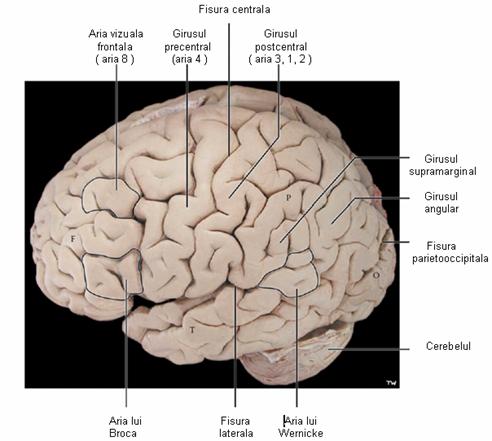 Figura 4
Figura 4
Structura emisferelor cerebrale (Figura 4) este reprezentata de substanta alba si substanta cenusie. Substanta alba la exterior, este alcatuita din trei tipuri de fibre :
de proiectie, ascendente (senzitive) si descendente (motorii), prin care cortexul este conectat cu celelalte structuri ale SNC;
comisurale, care fac legatura intre cele doua emisfere,conectandu-le la baza lor prin: comisura alba anterioara, corpul calos si trigonul cerebral.
de asociatie care fac legatura intre zone ale aceleasi emisfere.
Substanta cenusie formeaza la interior ganglionii bazali (corpii striati), iar la exterior, scoarta cerebrala.
Ganglionii bazali sunt mase de substanta cenusie, asezate la baza emisferelor cerebrale, lateral si deasupra talamusului, fiind reprezentati de nucleii caudat si lentiform. Nucleii bazali sunt pozitionati pe traseul cailor extrapiramidale, fiind conectati intre ei, dar si cu nucleii mezencefalici, diencefalici si cu cortexul cerebral. Nucleii bazali intervin in reglarea motilitatii automate, a comenzilor voluntare corticale,a tonusului muscular, pe care-l inhiba. Aceste celule formeaza un sistem complex de control, care coordoneaza activitatea musculara, ceea ce permite corpului sa indeplineasca diferite tipuri de miscari liber si inconstient. Acest tip de activitate musculara este implicat in balansarea bratelor in timpul mersului, in expresia fetei si in pozitionarea membrelor inainte de ridicarea in picioare sau de mers.
Cortexul cerebral (scoarța cerebrala) este un strat gros de aproximativ 3 mm de materie cenusie cu aspect cutat reprezentand suprafata exterioara a creierului. Aceasta parte a creierului a devenit atat de dezvoltata la oameni incat a necesitat plieri repetate pentru a avea loc in craniu. Depliata, ar acoperi o suprafata de 30 de ori mai mare.
Intre pliuri exista cateva santuri adanci, care impart fiecare din cele doua emisfere ale cortexului in patru zone numite lobi (Figura 5) Fiecare din acesti lobi indeplineste una sau mai multe functii specifice. Lobul temporal serveste pentru auz si miros, lobul parietal pentru pipait si gust, lobul occipital pentru vaz, iar cel frontal pentru miscare, vorbire si gandirea superioara.
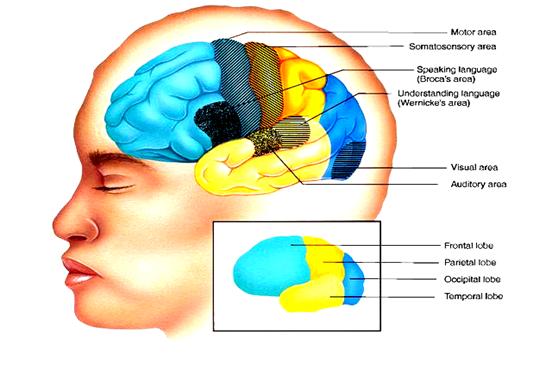
Figura 5
Din punct de vedere filogenetic si structural, aceasta prezinta doua zone: neocortexul si alocortexul. Neocortexul regiunea recent aparuta filogenetic, are cea mai mare dezvoltare la om fiind formata din 6 straturi de neuroni (2 straturi glanulare, 2 straturi piramidale, un strat molecular si unul fusiform).
Neocortexul din punct de vedere functional este impartit in:
neocortex receptor
neocortex motor
neocortex de asociatie
Neocortexul receptor reprezinta zona de proiectie corticala a sensibilitatilor specifice. Cuprinde arii senzitive si senzoriale
Neocortexul motor cuprinde arii (Figura 6) in care isi au originea fibrele piramidale si o parte a celor extrapiramidale.
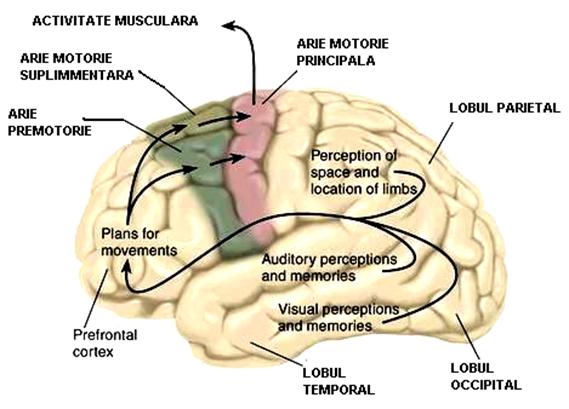
Figura 6
Neocortexul de asociatie (Figura 7) este o zona nou aparuta, extinsa in toate regiunile corticale, la nivelul careia excitatia nu produce manifestari senzitive sau motorii, rolul sau este de a asocia diversele arii corticale, indeplinind si functii psihice. Creierul uman are trei zone de asociatie: temporala (implicata in controlul activitatii vegetative si in determinarea personalitatii) prefrontala (cu rol in intelegerea cuvintelor, in memoria vizuala, in activitatea sexuala si in reactivitatea emotionala) parietooccipitala (cu rol in preluarea informatiilor primite de la aria somestezica si a informatiilor vizuale influentand declansarea unei activitati motorii corespunzatoare).
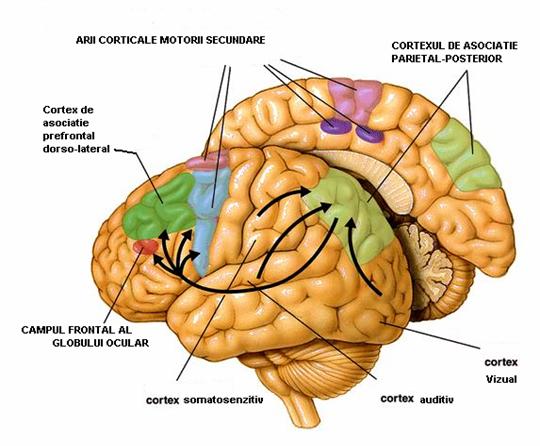 Figura 7
Figura 7
Alocortexul, sau zona veche a scoartei cerebrale este alcatuit din arhicortex (include bulbii olfactivi) si paleocortex (include hipocampul si o parte din hipotalamus). Se mai numeste si sistem limbic. Sistemul limbic are o structura mai simpla, numai din doua straturi celulare, si reprezinta sediul proceselor psihice afectiv-emotionale precum si a actelor de comportament instinctiv. Paleocortex are rol de integrare primaro-olfactiva, functie vegetativa si are legaturi cu glandele endocrine (hipofiza). Are rol in starile emotionale, instinctul alimentar si in procesul de invatare
I.2. Fiziologia Sitemului Nervos Central
Intreaga activitate a sistemului nervos se bazeaza pe impulsurile nervoase. Pentru a intelege mai bine si a ne forma o imagine cat mai apropiata de adevar, vom analiza structurile moleculare implicate (neuroni), fenomenele fiziologice care au loc (potential de actiune) precum si substantele chimice care actioneaza la acest nivel (mediatorii chimici).
I.2.1. Neuronul
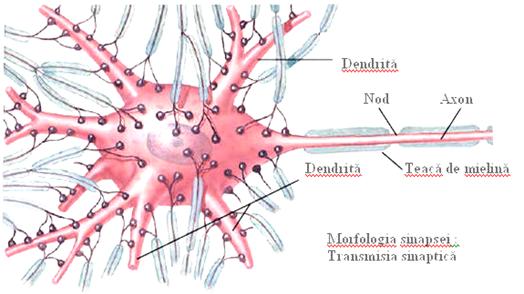
Unitatea morfofunctionala a sistemului nervos, este celula nervoasa sau neuronul (Figura 8). Este o celula specializata in generarea si conducerea unor semnale de natura electro-chimica numite impulsuri nervoase. Are forma ovala, rotunda, stelata, piramidala, si prezinta una, doua sau mai multe prelungiri (dendritele respectiv axonul), particularitate structurala ce ii confera functia de transmitere a impulsurilor nervoase. Un neuron este alcatuit dintr-un corp celular (soma) si prelungirile sale (dendritele si axonul).
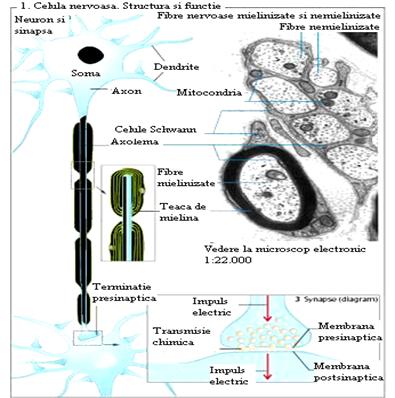 Figura
8
Figura
8
Corpul celular formeaza substanta cenusie din nevrax si ganglionii somatici si vegetativi. El este despartit de o membrana lipoproteica neurilema, are citoplasma-neuroplasma, ce contine organite citoplasmatice si un nucleu, obisnuit central, cu unul sau mai multi nucleoli. Sunt prezente organite celulare (mitocondrii, complex Golgi, reticul endoplasmatic, lizozomi) compusi intalniti si in alte celule, dar si organite specifice neuronului cum sunt corpusculii Nissl si neurofibrilele.
Corpusculii Nissl sunt constituiti din mase dense de reticul endoplasmatic rugos, la nivelul carora au loc sintezele proteice neuronale.
Neurofibrilele apar ca o retea omogena de fibre care traverseaza intreaga neuroplasma; au rol de sustinere si in transportul substantelor.
Dendritele sunt prelungiri celulare, uneori ramificate (ceea ce crește foarte mult posibilitatea contactului cu alte celule nervoase), care contin neurofibrile si corpusculi Nissl avand functia de a primi impulsuri si a le transmite spre corpul celular (conduc influxuri nervoase centripet sau aferent).
Axonul este o prelungire unica, lunga, alcatuit din axoplasma (continuarea neuroplasmei), in care se gasesc neurofibrile, mitocondrii si lizozomi, si este separat de o membrana, axolema, continuarea neurilemei. Axonul se ramifica in portiunea terminala, unele ramificatii fiind butonate (butoni terminali). Acestia contin, in afara de neurofibrile numeroase mitocondrii, precum si vezicule in care este stocata o substanta (mediator chimic). Axonii conduc impulsul nervos centrifug (eferent).
Fibra axonica este acoperita de mai multe teci. Teaca Schwann este formata din celule gliale, care inconjura axonul. Intre doua celule Schwann succesive se afla strangulatiile Ranvier (regiune nodala).
Majoritatea axonilor prezinta o teaca de mielina, secretata de celulele Schwann si depusa sub forma de lamele lipoproteice concentrice, albe, in jurul fibrei axonice (axoni mielinizati). Rolul tecii de mielina consta, atat in protectia si izolarea fibrei nervoase de fibrele invecinate, cat si in asigurarea nutritiei axonului.
Teaca Henle este o teaca continua, care insoteste ramificatiile axonice pana la terminarea lor, constituita din celule de tip conjunctiv, din fibre de colagen si reticulina, orientate intr-o retea fina care acopera celulele Schwann pe care le separa de tesutul conjunctiv din jurul fibrei nervoase. Aceasta teaca conjunctiva are rol nutritiv si de protectie.
Neuronul, asemanator celorlalte celule, prezinta o menbrana alcatuita dintr-un strat ce conține fosfolipide cu grupari nepolare hidrofobe in interior si grupari polare hidrofile in exterior.
Stratul lipidic este aproape impermeabil pentru apa si substantele hidrosolubile cum ar fi glucoza sau ionii, in schimb substantele hidrosolubile precum oxigenul si dioxidul de carbon il traverseaza cu usurinta. In acest strat lipidic se afla molecule de proteine cu rol de proteine de transport care actioneaza ca pompe ATP-aza Na / K si proteine ce joaca rolul de canale pasive pentru ioni care se deschid si se inchid in functie de schimbarile ce au loc in conformatia proteinei. Mai sunt de asemenea si proteine care au rol de receptori pentru fixarea neurotransmitatorilor si altele care functioneaza ca enzime (adenilat ciclaza) pentru catalizarea reactiilor la suprafata menbranei celulare.
In conditii de repaus (cand nu actioneza nici un stimul) neurilema este polarizata, fiind incarcata pozitiv pe fata externa si negativ pe fata interna, datorita impartirii inegale a Na+ si K+ de o parte si cealalta a menbranei.
In lichidul extracelular se afla o concentratie de Na+ mult mai mare comparativ cu celula care conține mai mulți ioni de K+.
Distributia clorului este asemanatoare cu cea a ionilor de Na , desi gradientul de concentratie este mic. Diferentele de ioni intre fata externa a membranei neuronale si interiorul celulei creeaza o diferenta de potential de aproximativ 70 mV, denumit si potential menbranar de repaus (se noteaza in mod conventional -70mV), mentinuta prin activitatea unor mecanisme menbranare, care funcționeaza cu consum energetic. In timpul potentialului de repaus menbrana este permeabila pentru K si impermeabila pentru Na
Intrucat potentialul membranar de repaus este de -70 mV, este de asteptat ca in repaus, in baza potentialului Nernst Na+ sa treaca in celula. Potentialul Nernst reprezinta potentialul electric de +61mV care se opune deplasarii ionului conform gradientului de concentratie. Ionii de Na+ nu patrund in celula intrucat canalele transmembranare pentru Na+ sunt inchise, iar cantitatile mici de ioni care traverseaza membrana sunt redistribuite prin interventia mecanismelor membranare.
Aceste mecanisme expulzeaza permanent Na+ patruns in celule si reintroduc K+, care iese lent si pasiv din celula datorita gradientului de concentratie dintre mediul celular si cel extracelular. Deoarece membrana neuronala este impermeabila pentru cei mai multi anioni din celula, iar efluxul K+ din celula este superior influxului de Na+, membrana se mentine in stare polarizata cu fata externa pozitiva fata de cea interna, negativa.
In urma aplicarii unui stimul cu intensitatea prag va avea loc depolarizarea membranei datorita cresterii permeabilitatii ei pentru ionii Na+ ca urmare a accentuarii influxului de Na+, polaritatea membranei se inverseaza, devenind negativa pe fata externa si pozitiva pe cea interna, diferenta de potential devine 35 mV si se numeste potential de.
Potentialul de actiune este de fapt schimbarea rapida a potentialului transmembranar urmata de o intoarcere la potentialul membranar de repaus.
Practic transmiterea depolarizarii de-a lungul fibrei nervoase reprezina influxul nervos (Figura 9).
Depolarizarea se propaga pas cu pas de-a lungul axonului, prin intermediul circuielor locale (fluxuri circulare sau curentii lui Herman). In axonii cu teaca de mielina, influxul nervos se propaga saltatoriu de la o strangulatie Ranvier la alta. In axonii fara teaca de mielina, influxul nervos se propaga intr-o singura directie, punctiform. Viteza de propagare a influxului nervos in axonii mielinizati este de 50 de ori mai mare decat in cei amielinici. Ulterior are loc repolarizarea care se declanseaza prin scaderea permeabilitatii membranei si a influxului pentru Na+, concomitent cresterii efluxului de K+, restabilindu-se astfel echilibrul ionic initial. Transportul ionic se realizeaza pasiv, gratie canalelor ionice (polipeptide sau lipopeptide cu greutate moleculara redusa) sau activ, prin intermediul pompelor ionice (agregate de proteine transportoare si enzime). Transmiterea unidirectionala a influxului nervos de la un neuron la altul, de la receptor la un neuron si de la neuron la efector se realizeaza prin intermediul unor structuri specializate numite sinapse.
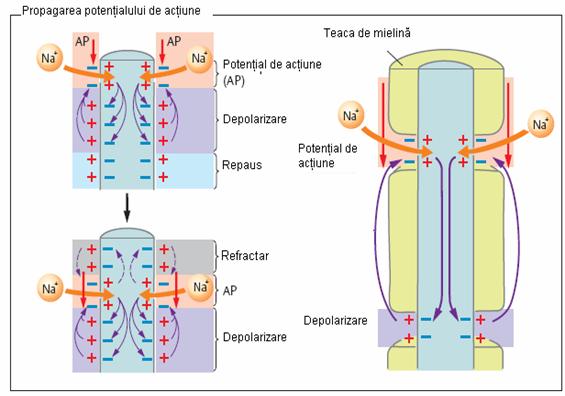 Figura 9
Figura 9
I.2.2. Sinapsa
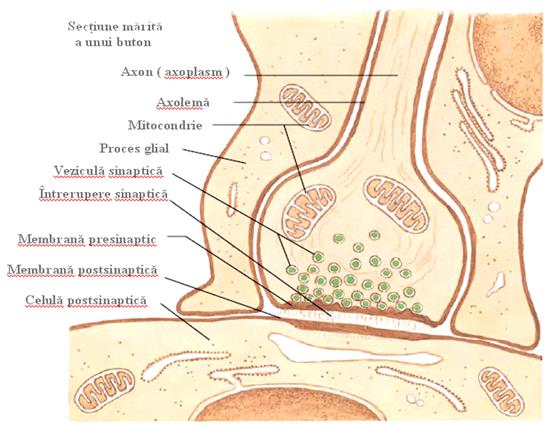
O sinapsa reprezinta locul unde membranele a doua celule excitabile sunt suficient de apropiate ca sa permita transmiterea informatiilor (Figura 10,11). Axonul unui neuron se termina (face sinapsa) cu dendritele unui alt neuron sau cu celula unui organ efector. La acest nivel exista diferentieri morfo-functionale ce determina excitatia sau inhibitia elementului postsinaptic atunci cand neuronul presinaptic intra in activitate. Transmiterea se realizeaza cel mai frecvent prin mediatori chimici (sinapse chimice) adica permite transmiterea impulsului numai de la o formatiune presinaptica (depozitul mediatorului) la formatiunea postsinaptica ce contine receptori specifici.
In cadrul sinapsei chimice, transmiterea sinaptica se realizeaza prin intermediul unei substante chimice specifice denumita neotransmitator chimic sau neuromeditator chimic si mai rar electric (sinapsa electrica).
Sinapsa chimica este alcatuita dintr-o regiune presinaptica si una postsinaptica separate printr-un spatiu sinaptic (fanta sinaptica).
Componenta presinaptica
Este reprezentata de terminatia butonata a axonului ce stabileste contactul sinaptic. Butonul terminal contine citoplasma, un numar variabil de mitocondrii, vezicule sinaptice si este limitat de o menbrana. Veziculele sinaptice reprezinta elementul caracteristic al regiunii presinaptice. Este o importanta modalitate de depozitare a substantelor mediatoare, ele deriva din reticulul endoplasmatic. Forma, dimensiunile si densitatea lor variaza in functie de mediatorul continut.
Fanta sinaptica
Reprezinta spatiul de 200 A cuprins intre cele 2 menbrane pre- si postsinaptica.La nivelul sau au fost identificate filamente cu orientare transversala ce asigura adezivitatea regiunii sinaptice si ar putea reprezenta un sistem de orientare pentru moleculele de mediator care il traverseaza.
Componenta postsinaptica
Este portiunea diferentiata a menbranei si a citoplasmei periferice a elementului postsinaptic (neuron, muschi, glanda). Menbrana postsinaptica este in general mai densa electronooptic si cuprinde structurile receptoare caracteristice mediatorului folosit.
 Figura
10
Figura
10
 Figura
11
Figura
11
Sinapsele cu transmitere chimica prezinta cateva caracteristici functionale:
unidirectionalitatea la nivelul sinapselor, mediatorul poate fi eliberat exclusiv din regiunea presinaptica care il contine si determina efectele sale doar la nivel postsinaptic unde exista receptori specifici. Acest lucru determina sensul de circulatie in intreaga retea neuronala si neuro-efectoare.
intarzierea sinaptica: etapele transimiterii chimice presusupune o latenta semnificativa. Datorita acestui fapt, intre momentul depolarizarii butonului terminal si momentul aparitiei activitatii postsinaptice se interpune un interval de 0,5-1,0 msec ce reduce in mod considerabil viteza de conducere in retelele multineuronale.
potentarea postetanica: cresterea frecventei de stimulare a terminatiei nervoase presinaptice determina o crestere a cantitatii de mediator eliberata in spatiul sinaptic la fiecare stimul.
fatigabilitatea stimularea prelungita si cu frecventa mare determina epuizarea rezervelor de mediator din butonul terminal si blocarea transmiterii sinaptice. In acest mod sinapsele actioneza ca un filtru ce nu lasa frecventele mari de stimulare sa progreseze in reteaua neuronala.
inexcitabilitatea electrica a membranei postsinaptice: mecanismele ionice ale potentialului de actiune nu pot fi declansate ca in restul membranelor excitabile prin stimulare electrica. Ele sunt conditionate de aparitia in zona a mediatorului chimic ce interactioneaza cu receptori.
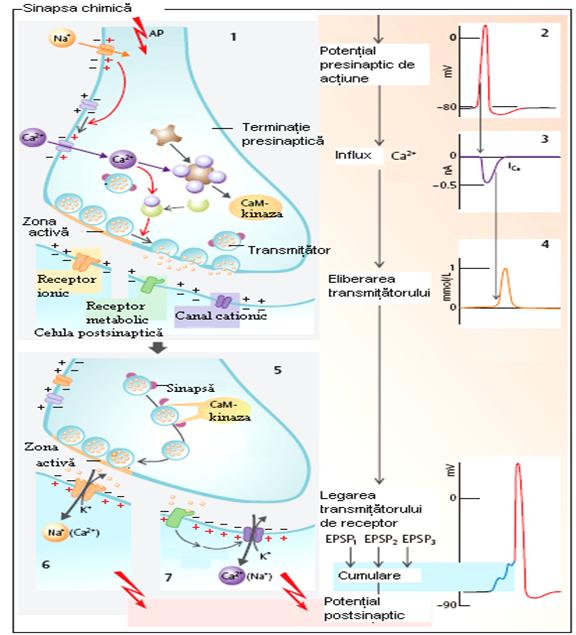
Transmia sinaptica (Figura 12) are loc dupa urmatoarele etape:
sinteza mediatorului
stocarea mediatorului
eliberarea mediatorului
traversarea spatiului sinaptic
actiunea postsinaptica a mediatorului
inactivarea mediatorului
Sinteza mediatorului consta in procesul biochimic de sinteza al substantei mediatoare, cu toate particularitatile specifice; are loc atat la nivelul somei neuronale cat si la nivelul terminatiei nervoase. Ambele zone de sinteza sunt prevazute cu echipamentul enzimatic necesar. Mediatorul chimic produs la distanta, in corpul neuronal, este transportat pana la nivelul terminatiei prin fluxul axonal ascendent.
Stocarea mediatorului este procesul prin care se creeaza rezervele presinaptice de substanta necesare pentru momentul in care un stimul presinaptic va provoca eliberarea de mediator intr-un ritm mult mai mare decat cel in sinteza. Sediul stocarii este reprezentat de veziculele sinaptice, restul de neuromediator aflat in exces fiind hidrolizat de enzime specifice AChE (acetilcolinestaraza) pentru acetilcolina, COMT (catechol-0-metiltransferaza) si MAO (monoaminoxidaza )pentru catecolamine. Se stie ca depozitele de mediator sunt compartimentate. Se descrie un compartiment stabil de depozit, ce cuprinde mediatorul de rezerva ce se elibereaza mai tarziu in cursul stimularii. Al doilea compartiment ar fi reprezentat de compartimentul labil, continand mediatorul imediat disponibil in momentul stimularii.
 Figura
12
Figura
12
Eliberarea mediatorului este actiunea prin care substanta chimica mediatoare ajunge in spatiul sinaptic. Acest proces este descris printr-un fenomen de secretie rapida si brusca declansat de aparitia potentialului de actiune la nivelul menbranei.
Depolarizarea menbranei butonului terminal determina pe langa patrunderea ionilor de Na si un influx masiv de ioni de Ca . Ionii de calciu din mediul extracelular patrund in oarecare masura prin canalele de Na voltaj-dependente, deschise de potentialul de actiune.
Cea mai mare parte din ionii de Ca patrund insa prin canale specifice de Ca voltaj-dependente, care se deschid cu o oarecare latenta. Acest influx de Ca ar constitui mecanismul de cuplare a potentialului de actiune cu secretia mediatorului. Ionii de Ca liberi din citoplasma determina modificari asupra mediatorului din depozitele imediat disponibile. Fixandu-se pe ATP din menbrana veziculara, ionul ar provoca schimbari de conformatie echivaland cu adeziunea veziculei de zona de eliberare, spargerea menbranei si contractie urmate de expulzia continutului in spatiul sinaptic. Dupa golirea continutului vezicular prin exocitoza, menbrana acesteia ar fi incorporata in structura menbranei presinaptice, din care ulterior s-ar forma noi vezicule care se incarca cu neuromediator.
Se considera ca eliberarea neuromediatorilor ar avea loc sub forma unor pachete moleculare egale intre ele, numite cuante. In repaus cuantele se elibereaza una cate una, cu frecventa scazuta.
In momentul aparitiei potentialului de actiune in teritoriul presinaptic, frecventa de descarcare a cuantelor de mediator ar creste asa mult incat prin sumare, ele provoaca aparitia potentialelor postsinaptice ce se pot propaga. O singura descarcare este echivalenta cu eliberarea simultana a peste 150 de cuante.
Traversarea spatiului sinaptic se desfasoara prin trecerea cuantelor eliberate prin acesta gratie miscarilor browniene tinzand sa ajunga la menbrana postsinaptica.
Actiunea postsinaptica a mediatorului: ajuns la nivelul menbranei postsinaptice, mediatorul isi manifesta actiunea prin cuplarea cu receptorii specifici.
Receptori inclusi in structura menbranei postsinaptice, reprezinta molecule a caror conformatie stereochimica le permite sa interactioneze specific cu molecula de mediator. Aparitia complexului mediator-receptor determina modificari in structura menbranei postsinaptice. Transformari reversibile ce duc la modificari de permeabilitate ce stau la baza raspunsului postsinaptic.
Inactivarea mediatorului consta in eliminarea mediatorului eliberat, in vederea reluarii ciclului la stimulul urmator. Viteza mare cu care are loc acest proces se datoreaza urmatoarelor mecanisme:
inactivarea postsinaptica realizata cu ajutorul enzimelor hidrolizante din menbrana postsinaptica. Aceste enzime aflate in apropierea receptorilor desfac complexul mediator-receptor pe masura ce acesta se formeaza.
captarea postsinanptica: o parte din mediatorul ce nu a fost fixat pe receptori este captat de structura postsinaptica si trecut in citoplasma acesteia, unde este inactivat.
difuziunea extrasinaptica: o parte din mediatorul eliberat difuzeaza in spatiul extracelular, unde este inactivat de enzimele hidrolizate cu sediu extracelular sau captat de celulele extrasinaptice
recaptarea :elementul nervos presinaptic capteaza o parte din mediatorul eliberat in vederea reutilizarii sale.
I.2.3. Neuromediatorii chimici
Neurotransmitatorii sunt mediatori chimici care sunt eliberati in fanta sinaptica drept raspuns la aparitia unui potential de actiune la nivelul terminatiei nervoase. Acestia pot fi excitatori sau inhibitori, in raport cu modificarile de configuratie proteica pe care le produc la nivelul membranei postsinaptice.
Prin interactiunea unui neurotransmitator excitator cu receptorul rezulta o crestere a permeabilitatii pentru cationi, in timp ce raspunsurile inhibitorii reflecta cresterea selectiva a permeabilitatii pentru ionii de clor. Un receptor postsinaptic poate fi excitat sau inhibat reflectand existenta ambelor tipuri de receptori in acelasi neuron postsinaptic. Mai mult decat atat, acelasi neurotransmitator poate fi inhibitor intr-o anumita zona si poate provoca excitatie in alta zona.
Substantele care indeplinesc criteriile de neurotransmitatori se impart in doua grupe: mediatori cu molecula mica si neuropeptide cu molecula mare.
Mediatorii cu molecula mica apartin unor clase diferite de substante chimice. Sintetizate si stocate in veziculele sinaptice din axonul terminal, ele actioneaza rapid asupra canalelor ionice de la nivelul membranei postsinaptice. Dintre neurotransmitatorii cu molecula mica fac parte: acetilcolina, noradrenalina, adrenalina, dopamina, serotonina, histamina, acidul gama-aminobutiric, glicina, glutamatul, aspartatul, oxidul nitric.
Mediatorii chimici de tipul neuropeptidelor nu sunt sintetizati in terminatiile presinaptice, ci in soma neuronala, o data cu sinteza de noi vezicule. Apoi acestea sunt transportate pana la capatul fibrei nervoase prin curentul axonal al axoplasmei, cu viteza foarte mica, de numai cativa centimetri pe zi. In cele din urma, aceste vezicule isi elibereaza continutul ca raspuns la potentialul de actiune in acelasi mod cu veziculele continand mediatori cu molecula mica.
Acidul gama-amino-butiric (GABA)
Este unul dintre neuromediatorii centrali inhibitori. Se gaseste in concentratii mari in substanta cenusie periapeductala, nucleul caudat, talamusul medial.
Sinteza are loc prin decarboxilarea acidului glutamic implicat in metabolismul oxidativ al hidrocarbonatilor la nivel de sistem nervos central.
Inactivarea are loc prin transaminare sub influenta GABA-transaminazei.
GABA determina efecte inhibitoare la nivelul scoartei, al nucelilor amigdalian si caudat. Eliberat de axonii celulelor Purkinje, determina inhibitia neuronilor din nucleul Deiters. Actioneaza la nivelul menbranei postsinaptice determinand hiperpolarizare si potentiale inhibitoare prin cresterea conductantei pentru ionii Cl.
Glicina
Este un transmitator mai putin comun, utilizat de neuronii maduvei spinarii, care inhiba muschii antagonisti. Distributia cerebro-spinala a glicinei este mai limitata decat a altor aminoacizi inhibitori. Cele mai mari concentratii de glicina se gasesc in trunchiul cerebral, in cerebel si in coarnele anterioare ale maduvei spinarii. Actiunea hiperpolarizanta post-sinaptica se realizeaza prin intermediul unui receptor cuplat la canalul de Cl. Actiunea canalului respectiv produsa de glicina provoaca, la fel ca in cazul receptorului gama-amino-butiric, cresterea permeabilitatii pentru ionul Cl.
Glutamatul
Este transmitator excitator major de la nivelul creierului. Aproape toate celulele cerebrale au receptori care ii raspund. Exista 4 tipuri de receptori pentru glutamat; ei raspund si controleaza canalele cationice cu conductanta mare care sunt permeabile la Ca , Na , K . Majoritatea neuronilor care secreta glutamat se gasesc la nivelul cortexului cerbral si in hipocamp.
Cantitatea excesiva de glutamat este toxica pentru neuroni, prin influxul mare de ioni de Ca intracelular. Neurotoxicitatea glutamatului se realizeaza in 2 faze sub influenta acumularii sale la nivelul extracelular.
Hipoxia, hipoglicemia, ischemia (lipsa aportului de sange) convulsiile prelungite si traumatismele cerebrale determina cresterea glutamatului extracelular. Aceste situatii provoaca intr-o prima faza umflarea si distensia neuronilor, insotite de cresterea influxului de Na , urmata in faza secunda de intrarea si acumularea excesiva de ioni de Ca
Alterarile menbranare si mitocondriale produse de acumularea excesiva de Ca provoaca fenomene de depolarizare prelungita si dereglari metabolice multiple intracelulare, ca urmare a activitatii enzimelor dependente de Ca de tipul fosfolipazei A. Excesul de radicali liberi si de derivati ai acidului arahidonic rezultati produce o excito-toxicitate care este ireversibila la nivelul principalelor ultrastructuri neuronale.
I.2.4. Sinapsa neuromusculara (placa motorie)
La acest nivel se produce contractia musculara. Acest proces se desfasoara in mai multe etape din momentul contractiei si pana la relaxarea finala. Initierea potentialului de actiune se realizeaza in muschiul normal "in situ", la nivelul placii motorii. Impulsul nervos determinand depolarizarea butonului terminatiei nervoase motorii, declanseaza influxul ionilor de Ca atat prin canalele de Na si mai ales prin canalele de Ca voltaj dependente de neurilema. In consecinta, se produce eliberarea masiva a acetilcolinei care traversand spatiul sinaptic, se cupleaza cu receptorii colinergi N din menbrana postsinaptica. Cuplarea cu subunitatea receptoare determina deschiderea canalului cationic din structura complexului si aparitia unei depolarizari locale. Cand aceasta depolarizare atinge pragul, se produce deschiderea canalelor ionice voltaj-dependente din sarcolema extrasinaptica si apare un potential de actiune similar cu cel din fibrele nervoase.
Cuplarea excitatie-contractie cuprinde ansamblul fenomenelor prin care potentialul de actiune initiat la nivelul sarcolemei determina cresterea brusca a concentratiei de Ca liber in sarcoplasma, necesara formarii puntii transversale si glisarii. Potentialul de actiune initiat la nivelul sarcolemei se propaga de-a lungul acesteia prin acelasi mecanism autoregenerativ exemplificat la nivelul membranei neuronale. Ajuns la nivelul tuburilor in T, potentialul se propaga de-a lungul triadelor descrise mai sus. La nivelul zonei de contact dintre tubul in T si cisternele RS s-au descris conexiuni si chiar canale.
In momentul depolarizarii tubului in T are loc un transfer de sarcina cu modificari de conformatie si deschiderea canalelor.
Sunt generati curenti ionici ce permeabilizeaza menbrana cisternei si determina eliberarea calciului depozitat.
Cresterea concentratiei Ca in sarcoplasma la o anumita valoare activeaza o pompa de Ca ATP-dependenta din menbrana cisternala, care readuce ionul in depozite.
Relaxarea musculara este asociata cu recaptarea Ca liber din sarcoplasma
|
Politica de confidentialitate |
| Copyright ©
2025 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |