
Fiziologia respiratiei
Respiratia reprezinta ansamblul fenomenelor fizice si chimice prin care se realizeaza schimbul de gaze intre organismul animal si mediul extern.
Respiratia este o functie vitala, intrucat doar cu mici exceptii viata este posibila numai in prezenta oxigenului.
La mamifere si om, respiratia se desfasoara in trei etape:
pulmonara, sau respiratia externa
sanguina
celulara, tisulara sau respiratia interna
Respiratia externa consta in introducerea aerului in pulmoni, unde oxigenul este retinut, iar bioxidul de carbon este eliminat; sediul acestor procese de schimb gazos sunt acinii pulmonari si capilarele adiacente lor: O2 trece din alveole in sange, iar CO2 trece din sange in alveole. Trecerea O2 din spatiul pulmonar in cel vascular poarta numele de hematoza.
Etapa sanguina incepe prin fixarea O2 pe hemoglobina, transportul lui sub forma de oxihemoglobinat de potasiu (HbO2K) la nivel tisular unde este cedat celulelor. CO2 este preluat la nivel tisular si transportat preponderent ca si bicarbonat la nivel pulmonar, unde este cedat alveolelor.
Respiratia interna reprezinta utilizarea O2 la nivel celular cu formarea de H2O, CO2 si energie; in mitocondrii, in procesul complex de la nivelul lantului de citocromi, hidrogenul activat cupleaza cu oxigenul, cu formarea de apa. CO2 rezulta prin decarboxilari.
1. Morfologia functionala a aparatului respirator
In procesul respiratiei participa sistemul respirator si o componenta vasculara, care se asociaza morfologic si se coordoneaza functional.
Sistemul respirator are doi poli: la polul pulmonar are loc respiratia externa, iar la polul tisular se desfasoara respiratia interna.
Polul pulmonar, reprezentat de aparatul respirator, prezinta o zona de conducere formata din caile aerifere si o zona respiratorie in care are loc schimbul gazos.
Caile aerifere se clasifica in extra pulmonare si intra pulmonare; ele nu participa la schimburile gazoase, dar prepara aerul pentru zona de schimb gazos.
Aerul patrunde in caile aerifere prin orificiile nazale. Mucoasa conductelor nazale este de tip respirator si olfactiv. Mucoasa respiratorie, bogat vascularizata, incalzeste aerul inspirat si il raceste pe cel expirat. Mucusul retine particulele inerte si purifica aerul. Acest proces de incalzire, purificare si saturare cu apa a aerului inspirat continua in trahee, bronhii si bronhiole.
Epiteliul mucoasei traheale prezinta celule ciliate in proportie de 80%; cilii au miscari in sens cranial, antrenand mucusul spre faringe. Atat celulele ciliate cat si cele secretoare de mucus scad numeric in bronhii si bronhiole.
In epiteliul traheal si bronhiolar mai sunt prezente celule secretoare de surfactant si celule cu secretie interna de: STH, TSH, ACTH, glucagon, gastrina, serotonina, histamina, catecolamine si prostaglandine.
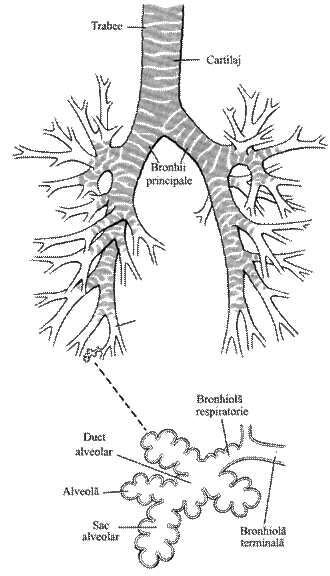 Peretele traheal
prezinta inele cartilaginoase incomplete, iar bronhiile mari au inele
cartilaginoase complete; pe masura ce diametrul bronhiolelor se
reduce, cartilajul se reduce treptat. La nivelul bronhiolelor terminale,
cartilajul este inlocuit de musculatura neteda dispusa circular;
aceste bronhiole conecteaza unitatile morfo-functionale ale
pulmonilor: acinii pulmonari (fig 1)
Peretele traheal
prezinta inele cartilaginoase incomplete, iar bronhiile mari au inele
cartilaginoase complete; pe masura ce diametrul bronhiolelor se
reduce, cartilajul se reduce treptat. La nivelul bronhiolelor terminale,
cartilajul este inlocuit de musculatura neteda dispusa circular;
aceste bronhiole conecteaza unitatile morfo-functionale ale
pulmonilor: acinii pulmonari (fig 1)
Acinul pulmonar are 3-5mm lungime si este format din 10-20 bronhiole respiratorii din care se desprind canale respiratorii care conecteaza sacii alveolari; acestia sunt formati din alveole cu diametru diferit (0,15-0,33mm).
Alveolele pulmonare au forma poliedrica in deflatie si forma hemisferica in inflatie. Suprafata lor totala insumeaza circa 100m2 la om. Peretele alveolar este acoperit in proportie de 50-70% din suprafata sa de capilarele pulmonare, impreuna formand suprafata de schimb gazos (fig 2).
Fata interna a alveolelor este captusita cu Fig 7.8 un film de surfactant care impiedica colabarea alveolelor mici in deflatie.
Vascularizatia pulmonilor este de tip nutritional si respirator. Vasele nutritionale provin din marea circulatie si se distribuie tesutului pulmonar pana la nivel de acini pulmonari. Vasele de tip respirator apartin micii circulatii si participa la procesul respirator; ele se distribuie acinilor pulmonari.
Inervatia pulmonilor este vegetativa; fibrele adrenergice, colinergice si purinergice se distribuie musculaturii netede a bronhiilor si a peretilor vasculari; fibrele nervoase sunt senzitive, bronho si vasomotorii, si bronhosecretorii.
Aparatul elastic pulmonar este o retea fina de fibre elastice perialveolare, care se continua cu o retea formata din fibre elastice mai groase peribronhial, care converg spre hilul pulmonar.
2. Mecanica respiratiei
Improspatare permanenta a aerului alveolar la schimb cu aer atmosferic constituie ventilatia pulmonara; aceasta se realizeaza prin variatiile de presiune intrapulmonare in decursul ciclurilor respiratorii. Un ciclu respirator se compune din: inspiratie, expiratie si pauza; numarul ciclurilor respiratorii pe minut constituie ritmul respirator. Acesta se modifica in raport cu necesarul de oxigen al tesuturilor.
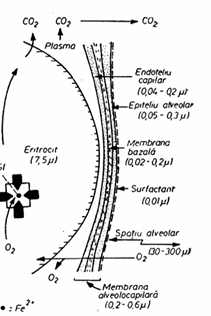
Fig 2. Bariera alveolo capilara
Presiune intrapulmonara a aerului se schimba datorita modificarii volumului cutiei toracice sub actiune muschilor respiratori. Din acest punct de vedere, muschiul diafragmatic este cel mai important: deplasarea cu un centimetru a diafragmei creste diametrul cranio-caudal al cutiei toracice si volumul sau cu valoarea ariei diafragmatice. Diametrele dorso-ventral si transversal ale cutiei toracice se modifica sub actiunea muschilor intercostali: externi sau inspiratori si interni sau expiratori .
In eupnee (respiratia linistita, de repaus) muschii expiratori sunt preponderent pasivi. Ei devin activi in expiratia fortata, alaturi de muschii abdominali. In dispnee (respiratia dificila) mai intervin muschii gatului, pectorali si abductori ai membrelor.
Tesutul pulmonar urmeaza cursa cutiei toracice determinand variatiile de presiune intrapulmonara. Aceasta se datoreaza functiei pleurei, care asigura aderenta functionala a tesutului pulmonar la cutia toracica.
3. Volumele pulmonare
Aerul prezent in aparatul respirator la un moment dat este format din spatiul mort total si aerul din acinii pulmonari.
Volumul de aer din spatiul mort total se imparte in volumul din spatiul mort anatomic cuprins in caile aerifere pana la nivel de bronhiole terminale si volumul de aer din spatiul mort fiziologic prezent in alveolele neperfuzate cu sange.
Volumul respirator curent este volumul de aer care intra si iese din pulmoni in respiratia eupneica; in medie, acest volum este de 10ml pe kg corp.
Volumul inspirator de rezerva este volumul de aer care intra in pulmoni dupa o inspiratie eupneica, prin inspiratie fortata.
Volumul expirator de rezerva este volumul de aer expirat fortat, dupa o expiratie eupneica.
Volumul rezidual este volumul de aer care ramane in pulmoni dupa o expiratie fortata.
Volumul de ventilatie alveolara este volumul de aer care intra si iese din acinii pulmonari si este singurul aer implicat in schimburile gazoase alveolare.
Capacitatea vitala a pulmonilor este volumul maxim de aer care poate intra si iesi din pulmoni.
Capacitatea pulmonara totala este suma volumelor de aer al capacitatii vitale si aerul rezidual (fig 3).
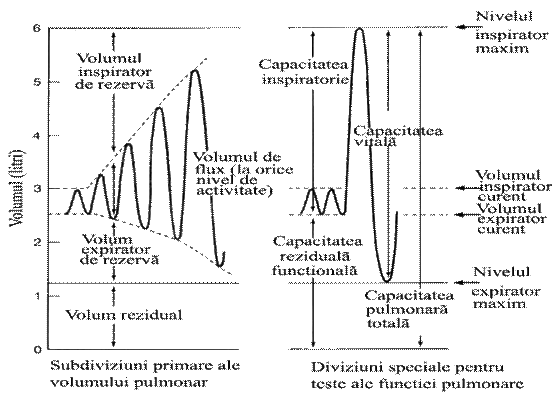
Fig 3. Volumele pulmonare
Coeficientul de ventilatie reprezinta proportia dintre aerul proaspat intrat in spatiul de schimb si aerul folosit. Circa doua treimi din aerul de schimb la sfarsitul inspiratiei este aer proaspat. Cu cat frecventa respiratiilor este mai mare, proportia de aer proaspat ajuns pe suprafata de schimb este mai mica. In polipnee, (respiratie cu frecventa mare si amplitudine mica) se ventileaza numai spatiul mort anatomic.
Problemele colabarii alveolelor. Existenta alveolelor cu diametre diferite in conditia realizarii unei tensiuni egale, in deflatie se creeaza probleme de ordin fizic, prin tendinta de colabare a alveolelor si a sacilor alveolari mici. Prin aceasta se reduce suprafata efectiva de schimb gazos.
Conform legii lui Laplace, diferenta de presiune dintre interiorul si exteriorul unui balon este proportionala cu 2T/R, in care T este tensiunea peretelui pe unitatea de lungime iar R este raza balonului. Daca doua baloane au aceeasi tensiune a peretelui, dar raza unuia este jumatate din a celuilalt, balonul mic va avea o presiune interna de doua ori mai mare; cand cele doua comunica intre ele, balonul mic se va goli in cel mare, iar peretii lui se colabeaza: P1=2T/R → P2=2T/(½R)=4T/R
O situatie similara exista in pulmoni, daca socotim alveolele si sacii alveolari ca niste baloane interconectate: alveolele mici tind sa se colabeze prin golirea aerului in alveolele si sacii alveolari mai mari.
Tensiunea peretelui alveolar depinde de proprietatile peretelui alveolar si de tensiunea superficiala la interfata cu aerul.
Surfactantul care captuseste alveolele este cel mai bun tensioactiv. El determina o foarte scazuta tensiune superficiala, care creste daca filmul de surfactant este intins si scade daca este comprimat. Intrucat surfactantul urmeaza peretele alveolar, la intindere creste tensiunea superficiala, compensand cresterea razei. Efectul este de a minimaliza diferentele de presiune dintre alveolele mari si mici, si de a reduce sansa colabarii, permitand inflatia oricarei alveole colabate.
Surfactantul este secretat de celulele alveolare si are un turnover de circa 12 ore.
Pierderile de apa si caldura prin respiratie
Aerul inspirat este incalzit si saturat cu apa in drumul sau spre acinii pulmonari. Prin aerul expirat se pierde atat caldura cat si apa. Cresterea ventilatiei pulmonare mareste proportional aceste pierderi.
La unele specii (in special cele de desert), mare parte din apa aerului expirat este recuperata la nivelul cavitatilor nazale; mucoasa respiratorie, bogat vascularizata, incalzeste aerul inspirat si il raceste pe cel expirat. Cu aceasta ocazie, vaporii de apa condenseaza si astfel apa este recuperata.
Unele mamifere si pasarile controleaza pierderile de caldura prin sistemul respirator, cu scopul reglarii temperaturii corpului. Cresterea pierderilor de caldura la mamifere se realizeaza prin respiratia orala si prin hiperventilatie sau polipneea termica, prin care se ventileaza doar spatiul mort anatomic.
Frecventa respiratiilor si tipuri de respiratie
Frecventa respiratiilor se masoara in numarul ciclurilor respiratorii pe minut. In eupnee, frecventa respiratiilor este diferita in functie de specie, talie si varsta animalelor (Tabelul 1).
|
Specia |
Frecventa |
Specia |
Frecventa |
|
Balena |
Cobai | ||
|
Vaca |
Porumbel | ||
|
Oaie |
Rata | ||
|
Porc |
Gaina | ||
|
Caine |
Gasca | ||
|
Pisica |
Curca | ||
|
Iepure |
Tabelul 1. Frecventa respiratiilor pe minut la animale
Cresterea frecventei se numeste tahipnee, iar reducerea ei bradipnee. Tahipneea fiziologica moderata se inregistreaza la femelele in calduri, in lactatie si gestatie, precum si in efort fizic.
Tipurile de respiratie sunt: tipul costal, la care predomina deplasarile peretilor costali (caine, caii din rasele usoare); tipul abdominal, la care ponderea o detin miscarile abdominale (rumegatoarele mari) si tipul diafragmatic (la iepure) cu predominarea miscarii diafragmului.
4. Schimburile gazoase
Schimburile de gaze, O2 si CO2 se realizeaza la cei doi poli ai sistemului respirator. La polul pulmonar schimbul reciproc de gaze se realizeaza intre aerul alveolar si capilarele pulmonare adiacente alveolelor. La polul tisular schimbul gazos se realizeaza intre sangele capilarelor sistemice si toate celulele corpului animal. Aceste schimburi se realizeaza prin difuziune pe baza gradientelor O2 si CO2 intre compartimentele de schimb. Acest proces este insotit de o serie de reactii chimice descrise de fenomenul Hamburger.
Masa de gaz transferata (M) depinde de aria disponibila pentru difuziune (A), distanta de difuziune (X), coeficientul de difuziune (D) si de diferenta de concentratie intre suprafetele de schimb (a1-a2): M=A∙D(a1-a2)/X.
Pentru facilitarea schimburilor, suprafata de schimb trebuie sa fie cat mai mare, iar distanta de difuziune sa fie cat mai mica.
Necesarul de O2 si productia de CO2 sunt dependente de masa corporala, iar schimbul de gaze este dependent de suprafetele de schimb specializate.
Rata de difuziune a unei substante este invers proportionala cu greutatea ei moleculara (Legea lui Graham). O2 si CO2 au molecule de marimi similare si vor difuza cu viteze similare, in conditiile unui gradient mai mare al O2 si o difuzibilitate mai mare a CO2. La nivel tisular O2 este utilizat, iar CO2 este produs aproximativ cu aceeasi rata. Raportul dintre O2 consumat si CO2 produs (numit cat respirator) variaza cu natura substratului metabolizat si are valori de 0,7; 0,8 si 1,0 pentru lipide, proteine si respectiv glucide. Ponderea glucidelor in hrana si respectiv proportia lor mai mare ca substrat metabolizat determina ca valoarea catului respirator sa tinda catre 1; in general numarul de molecule de O2 consumat este egal cu numarul de molecule de CO2 produse.
4.1. Schimburile gazoase alveolare
Difuziunea gazelor este determinata de presiunea lor; fiind in amestec, compozitia gazelor se exprima in presiuni partiale ale acestora in procente de volum. Presiunea partiala a O2 (PO2) in aerul atmosferic inspirat este de 20,9vol% iar PCO2 de 0,03vol%. In aerul expirat PO2 scade la 16,7vol% iar PCO2 creste la 3,8vol%.
PO2 in aerul alveolar este de 105mm Hg iar in sangele capilarelor pulmonare de 40mm Hg .
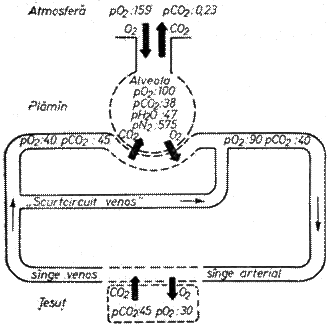
Fig 4. Schema schimburilor gazoase
Diferenta de 65mm Hg determina difuziunea O2 din alveole in capilare.
PCO2 in aerul alveolar este de 40mm Hg iar in capilarele sistemice de 46mm Hg; diferenta de 6mm Hg si difuzibilitatea mai mare a CO2 determina transferul CO2 din capilar spre alveola.
Ruta O2 este: aer alveolar, membrana alveolo-capilara, solubilizarea in plasma sangvina, trecerea prin membrana eritrocitara, fixarea pe hemoglobina
Ruta CO2: degajarea CO2 in plasma, solvirea in plasma, degajarea din plasma, traversarea peretelui alveolo-capilar, aer alveolar (fig 4.).
Procesele chimice care insotesc sau preced difuziunea O2 si respectiv CO2 sunt descrise de fenomenul Hamburger indirect:
O2 ajuns in eritrocit cupleaza hemoglobina redusa (HbH+) cu formarea oxihemoglobinei: O2+HbH+ → HbO2+ H+. Oxihemoglobina fiind un acid tare, cupleaza cu clorura de potasiu si formeaza oxihemoglobinatul de potasiu (HbO2K), forma cea mai importanta de transport a O2: HbO2+KCl → HbO2K+Cl-. Ionii de hidrogen cupleaza cu bicarbonatul de sodiu in plasma si formeaza acidul carbonic, care sub actiunea anhidrazei carbonice disociaza in CO2 si H2O:
H++NaHCO3 → HbCO3+Na+
H2CO3 → CO2+H2O
Na++Cl- → NaCl
In acest mod CO2 este eliberat din forma principala de transport (NaHCO3) si se permite difuzarea lui in alveola.
Relatia perfuzie-ventilatie pulmonara
Difuziunea gazelor respiratorii este dependenta, alaturi de presiunile partiale ale O2 si CO2 intre aerul alveolar si sangele capilar si de raporturile de suprafata de contact alveole-capilare, de rata de ventilatie si viteza de perfuzie a capilarelor pulmonare. Din punct de vedere fizic, difuziunea mai este dependenta de presiunea hidrostatica din arterele pulmonare, din atriul stang si de presiunea intraalveolara.
Datorita gravitatiei, in zona dorsala a pulmonilor presiunea intraalveolara este mai mare decat presiunea din arterele pulmonare, determinand colabarea partiala a capilarelor pulmonare; aceasta determina reducerea ratei de perfuzie, cu diminuarea difuziunii gazelor.
In zona mijlocie a pulmonilor, presiunea arteriala pulmonara este mai mare decat presiunea intraalveolara, realizand o perfuzie sanguina si de difuziune a gazelor adecvate.
4.2. Schimburile gazoase tisulare
In capilarele sistemice PO2 este de 95mm Hg, iar in celule de 40mm Hg. PCO2 in capilarele sistemice este de 40mm Hg iar in celule de 47mm Hg. Aceste diferente divizeaza CO2 din celule spre capilare. Procesele chimice care insotesc difuziunea gazelor sunt descrise de fenomenul Hamburger direct:
Atat in plasma, dar mai ales in eritrocite, CO2 sub actiunea anhidrazei carbonice formeaza acidul carbonic, care disociaza si determina scaderea pH-ului cu 0,2 unitati. Ionii de hidrogen cupleaza cu HbO2K eliberand oxigenul, care va difuza spre celule
CO2+H2O → H2CO3
H2CO3 → H++HCO3-
HbO2K+H+ → HbH+O2+K+
Ionul bicarbonic difuzeaza in plasma la schimb cu clorul si formeaza bicarbonatul de sodiu, crescand rezerva alcalina:
HCO3-+Na+ → NaHCO3
Cl-+K+ → KCl
La nivel celular, O2 este utilizat in mitocondrii; hidrogenul activat pe citocromi cupleaza cu O2 si formeaza apa. Reactia este exergonica, parte din energie fiind captata in legaturile macroergice de ATP. CO2 rezulta prin decarboxilari.
4.3. Functia respiratorie a sangelui
Legatura dintre cei doi poli ai sistemului respirator este realizata de sange. La nivel pulmonar O2 este preluat, transportat de sange si cedat tesuturilor, de unde este preluat CO2, adus la nivel pulmonar si cedat alveolelor. Cele doua gaze respiratorii, O2 si CO2 sunt transportate in combinatii reversibile si ca solviti in plasma.
4.3.1. Transportul oxigenului in sange
Oxigenul difuzat din alveole in sange dispune de doua forme de transport: dizolvat in sange si legat labil de hemoglobina.
Cantitatea de O2 dizolvata in sange este mica, dar are o importanta biologica mare, intrucat acesta constituie forma de O2 care este cedata tesuturilor. Cantitatea de O2 dizolvat in sange depinde de coeficientul de solubilitate si presiunea sa partiala si este invers proportionala cu temperatura (Legea lui Henry). Coeficientul de solubilitate al O2 in sange la 37sC este de 2,4mlO2/100ml sange la presiunea de O2 de 1 atmosfera. La PO2 alveolar de 100mm Hg numai 0,3mlO2/100ml sange se gaseste sub forma dizolvata. Cresterea de circa 70 de ori a continutului de O2 al sangelui fata de plasma este determinata de prezenta pigmentului respirator-hemoglobina. Vertebratele folosesc ca si pigment respirator circulant hemoglobina, iar ca pigment respirator de stocare, mioglobina (in muschi). Exceptie intre vertebrate fac unele specii de pesti arctici care nu poseda pigmenti respiratori si care compenseaza partial acest deficit prin cresterea volumului de sange si a debitului cardiac.
La nevertebrate, se gasesc pigmenti respiratori diferiti, care includ: hemeritrina si hemocianina, iar la unele grupuri si hemoglobina.
Hemoglobina continuta in eritrocite este formata din hem si globina si contine fier in forma feroasa (Fe2+). Prin legarea O2 de hemoglobina rezulta oxihemoglobina; in absenta O2 se utilizeaza termenul de deoxihemoglobina. O molecula de hemoglobina (Hb) contine patru hemi (Hm), hemul deoxigenat fixand o molecula de apa: Hm4(H2O)+4O2 ↔4HmO2+H2O.
Oxigenarea Hb este o reactie chimica labila; la un continut de 15g Hb la 100ml sange si o capacitate de legare de 1,38mlO2/g Hb continutul in O2/100ml sange oxigenat este de 20ml, valoare numita capacitatea de oxigenare a sangelui.
Gradul de saturare a hemoglobinei cu O2 este prin urmare dependent de PO2. Daca toate locurile de pe hemoglobina sunt ocupate cu O2 sangele este saturat 100% si continutul in O2 al sangelui este egal cu capacitatea lui oxiforetica; ea creste proportional cu continutul de hemoglobina in sange, continut care variaza cu specia de animale si cu individul. Dupa cum s-a vazut, incarcarea cu oxigen a hemoglobinei in conditii fiziologice nu atinge valoarea capacitatii oxiforetice. Pentru comparatie se utilizeaza termenul de procent de saturatie, care exprima continutul sangelui in O2 ca si procente din capacitatea oxiforetica.
Prin oxidarea Fe2+ la Fe3+ (forma ferica) ca urmare a cedarii unui electron se obtine methemoglobina, care nu leaga O2. Formarea de methemoglobina se produce normal in sange, dar eritrocitele dispun de o enzima, methemoglobin reductaza, care reduce methemoglobina la forma functionala, feroasa a fierului.
Anumiti compusi, cum sunt nitratii si cloratii determina fie oxidarea hemoglobinei, fie blocarea methemoglobin reductazei; ca urmare creste proportia de methemoglobina si se reduce transportul O2.
Afinitatea hemoglobinei pentru monoxidul de carbon (CO) este de 200 de ori mai mare decat pentru O2; CO va disloca O2 si va satura hemoglobina, cu formarea de carboxihemoglobina. Efectul acestei saturari este echivalent cu deprivarea de O2 a metabolismului oxidativ sau anoxia (fig 5.).
Curba de fixare sau disociere a O2 descrie relatia grafica intre procentul de saturatie cu O2 si PO2. curba este sigmoida; prin logaritmare ea devine liniara, punctul de saturare fiind exprimat matematic prin formula de echilibru O2-Hb a lui Hill:
Log(J/100-J)=log K+nlog P
in care J=% de saturatie; P=PO2; n= panta (ea indica numarul de subunitati hemice interactionate).
Intersectia J/100-J cu axa da log K; aceasta este constanta de echilibru O2-Hb.
Oxigenarea primului grup hemic favorizeaza oxigenarea celorlalte grupuri hemice, probabil prin schimbari in conformatia globinei.
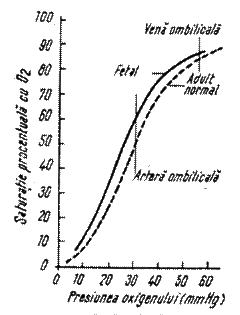
Fig 5. Curba de disociere a oxihemoglobinei F (linie intreaga) si a oxihemoglobinei A (linie intrerupta)
Afinitatea hemoglobinei pentru oxigen difera cu tipul de hemoglobina: embrionara, fetala, de adult. Hemoglobina fetala are o afinitate mai mare pentru O2 imbunatatind transferul de la mama la fat.
Hemoglobina cu mare afinitate pentru O2 nu va ceda oxigenul tesuturilor decat la o PO2 foarte scazuta; hemoglobina cu afinitate scazuta pentru O2 va facilita eliberarea O2 in tesuturi, mentinand o diferenta a PO2 mai mare intre sange si tesuturi, asigurand in acest fel o rata inalta a transferului de O2 spre tesut. Prin urmare, hemoglobina cu afinitate mare pentru O2 favorizeaza preluarea O2 de catre sange, iar cea cu afinitate scazuta faciliteaza eliberarea O2 in tesuturi. Functional, afinitatea hemoglobinei poate fi redusa in tesuturi si crescuta la nivel pulmonar.
Reducerea afinitatii hemoglobinei este data de: cresterea PCO2, scaderea pH-ului si cresterea temperaturii sangelui si a continutului sau in 2,3-difosfoglicerat(DPG).
Interactiunea dintre H+ si afinitatea hemoglobinei pentru O2 este descrisa de efectul Bohr: scaderea pH-ului sanguin reduce afinitatea pentru O2. Cresterea cantitatii de DPG la altitudine inalta (cu 10% la 3000m) reduce afinitatea hemoglobinei pentru O2 si favorizeaza oxigenarea tesuturilor. La pesti ATP-ul, iar la pasari inozitol fosfatul au efect mai mare decat DPG si au o concentratie mai mare in eritrocite, marind efectul Bohr.
Mioglobina contine un singur grup hemic, curba de disociatie fiind hiperbolica; ea nu este dependenta de pH).
 4.3.2. Transportul bioxidului de carbon in sange
4.3.2. Transportul bioxidului de carbon in sange
Din cantitatea de CO2 difuzata in sangele capilarelor sistemice, 8% se dizolva in plasma si 92% difuzeaza in eritrocite, unde in suita de reactii descrise de fenomenul Hamburger direct se furnizeaza HCO3-, care transferat in plasma formeaza NaHCO3; acesta reprezinta 65% din forma de transport a CO2.
In prezenta H+ si CO2 o parte din HbO2K disociaza cu formarea de HbH care se combina cu 27% din CO2 total prezent in sange, ca legatura carbamica a globinei (Gl): Gl+NH2+CO2 → Gl- Fig 5. Centrii respiratori NA-COO-+H2.
Gradul de formare a carbamatilor depinde de numarul gruparilor NH2 libere, de pH-ul sanguin si PCO2. la PO2 mica si PCO2 mare proportia legaturilor carbamice creste (efectul Haldane).
In eritrocite se formeaza mai multi compusi carbamici decat in plasma datorita unui numar mai mare de grupari NH2 libere, iar in plasma predomina HCO3.
Forma solubila de transport a CO2 este mai redusa decat in forma de carbamati, 2/3 din CO2 solubil fiind transportat in plasma si 1/3 in eritrocite.
5. Reglarea respiratiei
Cerintele de oxigen pentru activitatea metabolica variaza cu starea fiziologica a animalului si activitatile sale curente.
Asigurarea necesarului de O2 si CO2 si debarasarea adecvata de CO2 se realizeaza prin reglarea nervoasa si umorala a respiratiei externe.
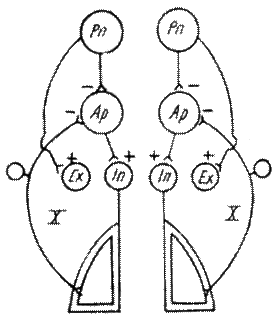
Respiratia eupneica este intretinuta de activitatea centrului inspirator bulbar, situat in portiunea ventro-laterala a nucleului tractului solitar; este alcatuit din neuroni care descarca potentiale de actiune pentru muschii inspiratori.
Un alt grup de neuroni, situati in nucleii ambiguu si retro ambiguu este constituit din neuroni inspiratori si expiratori.
In timpul respiratiei eupneice, centrul expirator bulbar actioneaza prin inhibarea activitatii centrului inspirator, determinand relaxarea muschilor inspiratori si consecutiv expiratia. Astfel, cei doi centri bulbari controleaza alternativ umplerea si golirea pulmonilor.In tesutul pulmonar exista receptori de presiune
care la intinderea tesutului pulmonar in inflatie genereaza, prin compresiune, impulsuri a caror frecventa este proportionala cu presiunea exercitata pe receptori. Aceste impulsuri sunt conduse in bulb, informand in permanenta despre gradul de inflatie al pulmonilor. Inspiratia este stopata cand nivelul de umplere cu aer al pulmonilor este la valoarea volumului respirator curent. Acest mecanism este adecvat pentru reglarea eupneei si se realizeaza pe baza reflexului de inflatie Hering-Brener.
Exista situatii cand necesarul de O2 creste si activitatea centrilor bulbari trebuie sa-si schimbe nivelul de activitate. Aceasta se realizeaza prin activitatea receptorilor localizati in arcul aortic si sinusul carotidian de la bifurcatia arterelor carotide interna si externa (fig 7.).
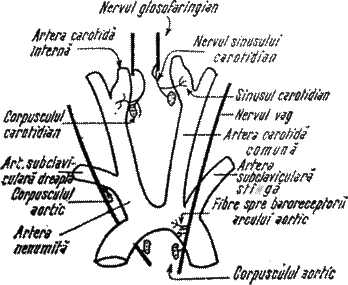
Fig 7. Ariile de receptie a schimbarii PO2
receptori sensibili la schimbarea PO2 in sangele arterial; ei determina schimbarea frecventei impulsurilor sosite la centrii nervosi, bulbul fiind informat de scaderea PO2 si in consecinta creste amplitudinea respiratiilor.
In efort creste si PCO2, care sensibilizeaza receptorii pentru O2 din sinusul carotidian. Prin aceasta, cresterea PCO2 potenteaza efectul scaderii PO2.
Receptorii sensibili la PO2 si PCO2 in lichidul cefalorahidian (LCR) se gasesc si pe planseul ventriculului IV in apropierea centrului inspirator. Variatiile de pH in sange sunt neimportante, dar absenta sistemelor tampon din LCR determina consecutiv cresterii PCO2, scaderea pH-ului care excita chemoreceptorii bulbari si creste amplitudinea respiratiei pana la corectarea PO2 si PCO2 la valori normale.
In zona rostrala a puntii se gaseste centrul pneumotoxic. Daca centrii bulbari sunt responsabili de amplitudinea respiratiilor, centrul pneumotoxic este responsabil de frecventa ciclurilor respiratorii.
Excitarea centrului pneumotoxic se realizeaza ca si a centrilor bulbari, prin modificarea PO2 si PCO2. centrul pneumotoxic excitat descarca impulsuri in centrul inspirator, inhiband inspirul si generand expirul. Activitatea centrului pneumotoxic poate fi reglata pentru limite largi de frecventa a ciclurilor respiratorii.
Baza morfo-functionala a centrului pneumotoxic este modul de sinaptizare interneuronala: cresterea numarului de sinapse, incetineste viteza de propagare a impulsurilor.
Scoarta cerebrala controleaza activitatea centrilor respiratori bulbo-pontini; emotiile produc modificari in ritmul respirator, dovedind controlul cortical al respiratiei. Modificarile voluntare ale ritmului respirator sunt posibile numai la om; apneea voluntara este posibila numai pana la pierderea cunostintei, dupa care ritmul respirator se reia.
Reflexe involuntare ca tusea, stranutul, deglutitia, modifica ritmul respirator.
La reptile si unele specii de pasari exista receptori sensibili la cresterea PCO2, care inhiba respiratia si le permite submersia pe perioade de timp mai lungi. Absenta acestor receptori la om si mamifere face imposibila submersia pe perioade mai lungi de timp, datorita declansarii inspirului chiar si in conditii de submersiune.
5.1. Reglarea bronhomotricitatii
Bronhiolele cu lumen mic, lipsite de cartilaj, dispun de musculatura neteda dispusa circular; aceasta este inervata vegetativ de fibre adrenergice, colinergice si purinergice. Astfel de fibre se distribuie si musculaturii netede arteriale adiacente.
Fibrele adrenergice sunt bronhodilatatoare (receptori β) si bronhoconstrictoare (receptori α).
Fibrele colinergice sunt bronhoconstrictoare (receptori muscarinici); actiunea acetilcolinei este potentata de serotonina si inhibata de noradrenalina si prostaglandina E2. Atropina are actiune bronhodilatatoare.
Reglarea neuroreflexa a tonusului musculaturii bronhice este predominant vagala.
Fibrele musculare netede bronhiolare prezinta receptori membranari pentru aminele biogene (histamina, serotonina) pentru prostaglandine si hormoni circulanti.
Factorii reglarii umorale actioneaza sinergic cu cei nervosi; catecolaminele produc bronhodilatatie si hiposecretie bronhica. Histamina, serotonina, bradichinina si prostaglandina F2 α produc bronhoconstrictie si hipersecretie bronhiolara.
6. Particularitatile respiratiei la pasari
Transferul gazelor respiratorii la pasari se realizeaza prin mici capilare aerifere cu sectiunea de circa 10μm, (fig 8) care se desprind din parabronhii; acestea reprezinta echivalentul sacilor alveolari de la mamifere.

Fig 8 Schema capilarelor aerifere la pasari
Parabronhiile sunt o serie de tuburi mici dispuse intre dorsobronhii si ventrobronhii, care au lumenul mai larg si care comunica cu mezobronhia; aceasta se leaga cranial de trahee. Acest ansamblu formeaza pulmonii la pasari (fig 9.).
In timpul respiratiei, volumul pulmonilor si al cutiei toracice se schimba putin, datorita prezentei sacilor aerieni conectati la pulmoni..
Cand acesti saci (caudal si cranial) sunt contractati, aerul este fortat in parabronhii. Modificari importante de volum se produc in timpul respiratiei numai in sacii aerieni toracic (cranial) si abdominal (caudal).
Aerul circula in doua directii in mezobronhii, dar unidirectional in parabronhii. In timpul inspiratiei aerul din trahee trece prin mezobronhii in sacii abdominali si prin ventrobronhii in sacii craniali. Din ventrobronhii si dorsobronhii in etapa urmatoare, aerul intra in parabronhii si canalele aerifere.
In timpul expiratiei aerul paraseste sacii abdominali, trece prin parabronhii si in masura mai mica prin mezobronhii in trahee.
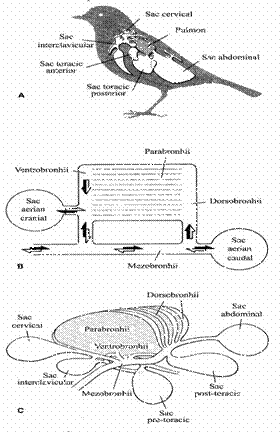
Fig 9. Schema aparatului respirator la pasari
Sacii aerieni craniali, al caror volum se schimba mai putin decat sacii caudali, cedeaza aerul ventrobronhiilor, apoi mezobronhiilor si traheei.
In acest fel se asigura un sens unidirectional al aerului prin parabronhii, atat in inspiratie cat si in expiratie, asigurand o ventilatie a zonelor de schimb gazos mai buna decat la mamifere.
|
Politica de confidentialitate |
| Copyright ©
2025 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |