
|
Ethanol fermentation technology - Zymomonas mobilis |
P. Gunasekaran and K. Chandra Raj Department of Microbial Technology, School of Biological Sciences, Madurai Kamaraj University, Madurai 625 021, India
Datorita diminuarii resurselor de combustubili fosili, productia microbiana de biocombustibili a cunoscut o crestere seminificativa in ultimii ani. S-a considerat ca etanolul va fi un combustibil alternativ in viitor. Chiar daca unele organisme, printre care si Clostridium sp., au fost considerate din punct de vedere etanologic microbi, drojdia Saccharomyces cerevisiae si bacteria facultativa Zymomonas mobilis sunt pretendenti mai buni pentru productia industriala de etanol. Zymomonas poseda avantaje fata de Saccharomyces in ceea ce priveste productivitatea in etanol si toleranta, ceea ce a incurajat cercetatorii sa exploateze abilitatea ei de a utiliza zaharoza, glucoza si fructoza prin caile Entner-Deudoroff. Obstacolele pentru Zymomonas sunt:
- imposibilitatea de a converti carbohidrati complexi precum celuloza, hemiceluloza si amidon la etanol;
- aparitia de bioprodusi ca si sorbitolul, acetoina, glicerolul su acidul acetic;
- formarea de polimeri extracelulari levan.
Pentru a imbunatati aceste neajunsuri, manipularea genetica a bacteriei Z. s=a indreptat spre extinderea gamei utilizabile a lui Z., de exemplu, genele responsabile de codificarea a catorva enzime hidrolitice de la specii bacteriene inrudite au fost clonate si transferate la Z. Interesant, un operon pet, (productia de etanol), a fost obtinut prin combinarea genelor pdc (piruvat decarboxilazei) si adhl (alcooldehidrogenazei) de la Z. si transferate la alte tulpini bacteriene pentru a le face ethanologenic novel strains. Prin mutatii clasice si abordare selectiva au fost obtinuti mutantii de Z. mobilis cu caracteristici fermentative imbunatatite si fara bioprodusi secundari. In plus fata de etanol, Z. mobilis a fost modificata metabolic pentru a produce L-alanina si L-acid lactic. Genele care codificau sinteza b-carotenilor au fost clonate si exprimate cu succes la Z. m. pentru a mari the fermented nutrients of farm animals .Aplicatiile levan in industria alimentara si farmaceutica asigura oportunitatea de a exploata Z. pe scara larga productia levanilor. Meritele lui Z. sugereaza posibila utilizare a microorganismului in productia industriala a diferitelor produse de fermenatie.
Resursele naturale de energie precum combustibilii fosili, (petrolul si carbunii) sunt utilizate cu o rata ridicata si acestea sunt estimate sa dureze putin timp. Astfel, sursele alternative de energie precum etanolul, metanul si hidrogenul sunt luate in considerare. Unele procese biologice pot oferi cai de producere a etanolului si metanului in cantitati mari. Interesul de a folosi biocombustibili ca si sursa de energie a dus la realizarea de studii care sa arate care este costul si eficienta proceselor industriale de obtinere a etanolului. S=au facut cercetari intense in obtinerea de organisme fermentative eficiente, substraturi fermentative cu cost redus , si conditii optime pentru ca fermentatia sa aiba loc. Traditional, etanolul a fost obtinut in loturi de fermentatie cu tulpini de drojdii care nu pot tolera o concentratie ridicata de etanol. Aceasta a necesitat imbunatatirea tehnologiei pentru obtinerea drojdiilor rezistente la concentratii ridicate in etanol. Z. mobilis, o bacterie gram negativa, este considerata ca un microorganism alternativ in vederea obtinerii etanolului ca si combustibil pe scara larga. Comparative laboratory- and pilot-scale studies on kinetics in loturi de fermentatie pentru Z. versus o varietate de drojdii au indicat faptul ca mai potrivit este Zimomonas fata de drojdii, avand avantajele urmatoare:
its higher sugar uptake and ethanol yield;
are o productie de biomas mai scazuta;
prezinta o toleranta mai ridicata fata de etanol;
nu necesita aditie controlata a oxigenului pe parcursul fermentarii;
posibilitatea de manipulare genetica.
Singurele limitari ale lui Z comparativ cu drojdiile sunt acelea ca substraturile utilizabile sunt doar glucoza, fructoza si zaharoza. Z. mobilis a fost prima data izolata din sursele de bauturi alcoolice precum palmierul de vin african, the Mexican 'pulque', si ca si contaminant al vinului de mere si al berii in tarile europene. Folosind o abordare taxonomica moderna, genul Z.[2] are o singura specie cu doua subspecii , Z. mobilis ssp. Mobilis si Z. mobilis, ssp. Pomaceae. Abilitatea de a utiliza zaharoza ca si sursa de carbon distinge Z. mobilis de Z. anaerobia . Este una din putinele bacterii facultative anaerobe care metabolizeaza glucoza si fructoza prin caile E - D, specifice microororganismelor aerobe . In conditii de anaerobioza, Zymomonas duce la produsi secundari precum acetoina, glicerol, acetat, lactat, care apar in productia redusa a etanolului din glucoza. Pe parcursul dezvoltarii Z. mobilis in fructoza, producerea de acetoina, acid acetic, acetaldehida a fost mai pronuntata decat in glucoza. Cu toate acestea, randamentul in celule a fost redus in timpul dezvoltarii in fructoza. In plus fata de etanoluk de fermentare, Z. mobilis prezinta posibile aplicatii in productia polimerilor. Levan, un polimer al unitatilor de fructoza unite prin legaturi b 2,6-fructosil, e produs de Z. in timpul dezvoltarii sale pe mediile cu zaharoza. Levanii microbieni nu prezinta importanta comerciala si sunt folositi ca si agenti de ingrosare, supendare si gelling. In ultimii ani, s=a incercat imbunatatirea strategiilor de producere a acestor levani.
BIOCHIMIA LUI ZYMOMONAS MOBILIS
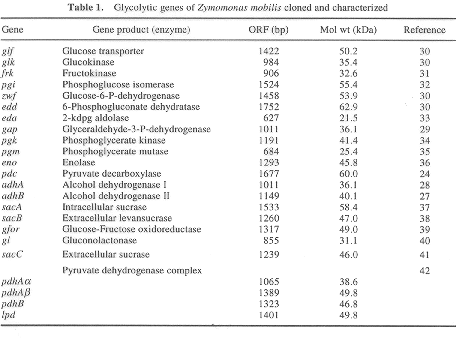
In Z. mobilis, D-glucoza si D-fructoza sunt transportate prin difuzie facilitata[5].Solutiile concentrate de glucoza nu inhiba enzimele cailor metabolice E - D, de cand se realizeaza conversia glucozei la etanol de catre acest microorganism rapid . Totusi, presiunea osmotica extracelulara a solutiei de glucoza poate fi rapid echilibrata astfel incat sa corespunda concentratiei interne de zaharuri. Concentratia ridicata de zaharuri scade potentialul total al apei, si exercita o presiune osmotica ce poate fi comparata cu aceea a unei solutii saline tari. Toleranta scazuta fata de saruri a lui Z. a aratat ca glucozo-6-fosfat dehidrogenaza si fosfogliceromutaza sunt enzime limitative, si fosfofructokinaza nu participa in caile metabolice E - D ale lui Zymomonas mobilis . Piruvat decarboxilaza(pdc) si alcool dehidrogenaza (Adh) sunt enzimele cheie in formarea etanolului. Pdc este o enzima unica la Zymomonas mobilis ce necesita tiaminpirofosfat pentru a-si desfasura activitatea( 8 Dawes, E. A., Ribbons, D. W. and Large, P. J., Biochem. J., 1966, 98, 795-803). In timp ce Adh I e zinc dependenta si asemanatoare cu Adh IV de la drojdii, Adh II e very unusual sa contina fier si nu zinc 9(Wills, C., Kratofil, P., Londo, D. and Martin, T., Arch. Biochem. Biophys., 1981, 210, 775-785 Adh II vine sa faciliteze continuarea fermentatiei la concentratii ridicate de etanol. SE VA ATASA FIGURA 1( Genele glicolitice clonate si caracterizate ale lui Zymomonas mobilis). La Zymomonas mobilis enzimele caii metabolice E - D sunt mai tolerante fata de etanol, decat the cell-free system ale Z. mobilis pot consuma rapid glucoza si sa produca etanol mai mult de 15% w/v 10 (Algar, E. M. and Scopes, R. K., J. Bacteriol., 1985, 2, 275-287. Membrana celulara de la Zymomonas si-a modificat continutul in acizi grasi pentru a nu suferi efecte secundare fata de actiunea etanolului. Majoritatea acizilor grasi de la Z. sunt acid miristic, acid palmitic si acid cis-vaccinic. Printre fosfolipide, fosfotidiletanolamina este in cantitatea cea mai mare. Concentratia ridicata de acid cis-vaccinic si unusual hopanoids din membrana asigura toleranta ridicata fata de etanol 11 (Buchholz, S. E., Dooley, M. M. and Eveleigh, D. E., Trends. Biotechnol., 1987, 5, 199-204). NADH- si NADPH-oxidaza sunt localizate in membrana celulara a lui Z. si catalizeaza oxidarea NAD(P)H. 12 (Pankova, L. M., Shvinka, Y. E., Beker, M. E. and Salva, E. E., Microbiologia, 1985, 54, 141-145). In plus, sunt prezente si catalazele enzimatice, superoxiddismutazele si peroxidazele. Aici, transferul de electroni pe calea lantului respirator nu e cuplat fosforilarea oxidativa. Si glucozo-fructozo oxidoreductazele si glucozodehidrogenazele duc la formarea acidului gluconic, care intra dupa fosforilare in caile metabolice E - D, dupa formarea NAD(P)H. Un mol din coenzima redusa formata e utilizata direct de manitol dehidrogenaza si NAD(P)H oxidaza, trecand in bioprodusi ca acetoina, acid acetic si acetaldehida. Productia de acetaldehida realizata de Zymomonas are rolul de a mari activitatea NADH oxidazei si de a micsora disponibilitatea NADH de a reduce acetaldehida la etanol cu Adh. Mutantii Adh ce deriva de la Zymomonas prezinta nivele ridicate ale productiei de acetaldehida izolate prin intermediul alcolului alilic ca si agent selectiv 13 (Wecker, M. S. A. and Zall, R. R., Appl. Microbiol. Biotechnol., 1987, 53, 2815-2820).
Zymomonas mobilis poseda atat fosfataza alcaline cat si acide. In timp ce activitatea fosfatazelor acide e mai ridicata in prezenta ionilor de Mg2+, fosfatazele alcaline necesita Zn (II) (Pond, J. L., Eddy, C. K., Mac Kenzie, K. F., Conway, T., Borecky, D. J. and Ingram, L. O., J. Bacteriol., 1989, 171, 767-774). Fosfatazele alcaline ale lui Zymomonas sunt asociate cu membranele si apar sub 2 izoforme.
Activitatea hidrolitica a zaharurilor e stimulata de zaharoza si fructoza. Rata hidrolitica a zaharozei si rata transfructozilarii sunt mai mari decat rata captarii de zaharuri a lui Zymomonas m. In timpul fermentarii zaharozei cu sub actiunea lui Z. au loc 3 tipuri de transfructozilari formandu-se 3 fructoze, oligozaharide , si polimeri mai mari ai fructozei si levani unde apa, zaharoza, si levani ai polifructozei respective pot actiona ca si acceptori.
Levanzaharoza a fost purificata din celle precum si din culturi de bulion. Trei enzime diferite zaharolitice au fost identificate la Z. mobilis: o zaharaza endocelulara, o levanzaharaza exocelulara si o zaharaza exocelulara. Aceste enzime sunt b-D-fructofuranozil-transferaze si sunt, prin urmare, capabile de a hidroliza zaharoza. Levanzaharazele sintetizeaza levani si poate fi la randul ei partial hidrolizata. Zaharazele si levanzaharazele nu hidrolizeza inulina, un plomer al fructozei cu masa molara mai mica.
Biologia moleculara a lui Z. mobilis
Marimea genomului tulpinii de Z, mobilis este de ordinul 1,53 plus minus 0,19 x 1019 Da, cam 56% din genomul E. coli si poate "acomoda"cam 1500 de cistroni. Compozitia bazelor AND pt. Z. mobilis (48,5 plus m. 1% G + C) a fost determinata prin denaturare termica.
Gena piruvatdecarboxilazei (pdc) a fost clonata de la tulpinile de Z. mobilis ATC 31821 si ATCC 29191. Regiunea codificatoare este de 1,7 kb lungime si codifica o polipeptida formata din 567 aminoazici cu o subunitate de masa de 60.8 kDa. Cum regiunea promotor nu contine secvente homoloage promotorului generalizat structurii E.coli, promotorul pdc nu e recunoscut de E. coli, cu toate ca gena clonata este exprimata relativ la nivele inalte sub controlul promotorilor alternativi.Compararea secventelor nucleotidice ale genei pdc din tulpina de Z. mobilis ATCC 29191 si ATCC 31821 arata existenta polimorfismului in izolari diferite.
Gena adh de la Z. mobilis a fost clonata folosind o placa indicatoare tehnica in care amestecurile de para rosanilina si de bisulfit sunt incorporate. Secventa AND din aceste gene contine un 'cadru deschis de lectura' open reading frame (ORF) care codifica o polipeptida formata din 383 de aminoacizi cu masa molara de 40 kDa. Gena adhl este transcrisa la un nivel scazut in E. coli de la promotorul P2 a lui Z. mobilis, dar este bine exprimata in E. coli sub controlul promotorului lac. Aceasta AdhI este responsabila de expunerea unei mici homologii cu alti produsi ai genei adh, dar expune o puternica homologie cu Adh IV de la S. cerevisiae.
Gena codificatoare pentru glicerinaldehid-3-fosfat dehidrogenaza (gap) a fost izolata la Z. mobilis completand o deficienta a tulpinii de E.coli. ORF pentru aceasta gena codifica 337 aminoacizi cu o masa moleculara de 36,1 kDa. Cu toate ca secventa gap a primului aminoacid are o homologie functionala considerabila si identitate cu aceeasi enzima de la alte eucariote si bacterii, de pare ca este mai inrudit de bacteriile termofile si de izoenzimele din cloroplate. Compararea aceste gene cu alte gene glicolitice de la Z. mobilis dezvaluie cateva trasaturi comune ale structurii genei, inclusiv un tipar conservat a PREJUDECATII CODONILOR (codon bias). Alte gene glicolitice care au fost clonate si caracterizate sunt redate in Tabelul 1.
Gnele codificatoare de la Z. mobilis pentru fosfogliceratkinaza (pgk) au fost clonate la E. coli si au fost secventate. Acest promotor-lipsa al genei pgk este localizat cu 225 perechi de baze in aval fata de decalajul de gene, si este parte a operonului gap.Secventa de aminoacizi dedusa din gena pgk a lui Z. mobilis e mai putin conservata decat alte secvente cunoscute ptr. Pgk, si, de asemenea, prezinta un grad ridicat de conservare a
structurii. In general, pozitia aminoacidului la granitele unei foi si a unei regiuni de helix b, si cele care realizeaza conexiunea in aceste regiuni e mai conservata decat aminocacizii pozitionati intre regiunile structurii secundare.
Gena phoC ptr. Z. mobilis codifica ptr . o fosfataza acida cu masa molara de 29 kDA. Promotorul sau cuprinde o caseta de regiune 35 pho, similara cu genele E. coli , precum si secventele regulatoare de la fostaza acida de la S. cerevisiae (pho5). Gena phoC contine un capat C 5 terminal bogat in AT, are un situs de legare ribosomala slabuta, si are o utilizare scazuta a biased codon decat genele puternic exprimate ale lui Z. mobilis. O comparatie a acestor gene arata ca promotorii tuturor acestor gene sunt similari in gradul de conservare al spatierii si identitatii cu genele in consesn propuse de Z. mobilis.
5 gene trp de la Z. mobilis au fost observate pentru abilitatea lor de a fi complementare cu mutantii trp. Organizarea genelor trp la Z. mobilis este similara cu aceea descoperita la speciile Rhizobium, Acinobacter calcoaceticus si Pseudomonas acidoovorans. Genele trpF, trpB, trpA apar ca fiind legate, dar ele nu sunt strans asociate cu genele trpD sau trpC in Z. mobilis.
Gena sacA codificatoare a unei zaharaze intracelulara de la Z. mobilis a fost clonata, secventata si caracaterizata. Proteina SacA este un monomer cu o masa molara de 58 kDa si secventa ei de aminoacizi prezinta o stransa homologie cu zaharaza intracelulara de la B. subtilis si invertaza drojdiilor. SacB codificand levanzaharaze extracelulare si sacC codificand zaharaze extracelulare au fost clonate si exprimate in E.coli. Analiza secventelor nucleotidice a genelor sacB descopera o ORF 1269 oerechi de baza long encoding pentru o proteina cu masa molara de 46,7 kDa. Secventa dedusa de aminoacizi a fost identica cu a secventei N-terminale cu cea dedusa din gena sacB de la Z. mobilis Z6C. Secventa de aminoacizi din SacB prezinta o similaritate scazuta cu cele din zaharazele cunoscute, dar foarte similara cu a levanzaharazei din Z. mobilis (61,5%), Erwinia amylovora (40,2%) si Bacillus subtilis (25,6%). Analiza secventei nucleotidice a sacC descopera un ORF de 1239 perechi de baze codificand o proteina de 46 kDa. Primii 30 de aminoaacizi dedusi din ORF au fost identici cu cei din secventa N-terminala a zaharazei extracelulare din tulpina de Z. mobilis ZM4. Aceasta gena sacC e localizata la 155 perechi de baze in aval de genele sacB formand o zaharaza de gene cluster. SacB si SacC sunt 2 enzime extracelulare secventate ale lui Z. mobilis care nu poseda secevente semnal.
Studiile au demonstrat cerintele secventei pentru localizarea membranelor sau transportul proteinelor in Z. mobilis. Fuziunea lacZ, care contine secvente ancora permitand asociatia membranara, e folosita pentru a izola AND-ul din Z. mobilis continand activitatea promotorului si secventele aminoterminale. Compararea acestor secvente si transcrierea site-urilor de initiere arata ca atat E.coli cat si Z. mobilis recunosc regiuni similare ale ADN pentru initierea transcrierii. 5-8 aminoacizi hidrofobi consecutivi in capatul amino terminal servesc drept ancora acestei proteine hibride in membrana E.coli si Z. mobilis.
Bioprocese potentiale:producerea etanolului
Producerea etanolului sub actiunea Z. mobilis a fost restrictionata la subtraturi ca glucoza, fructoza si zaharoza. Alternativ, substraturile de zahar brut precum sfecla de zahar, BAGASES si melasa au fost prezentate ca si substraturi pentru fermentatia directa la etanol. Tehnologia ADN recombinat a putut exploata si obtine tulpini microbiene pentru a obtine etanol din aceste substraturi agricole ieftine. Pentru a obtine tulpini etanologice utilizand substraturile de mai sus, fiecare Z. mobilis a fost transformata cu genele interesului dobandit de la alte organisme sau gene de la Z. mobilis implicate in sinteza etanolului au fost transferate la alte organisme cu caracteristicile cerute.
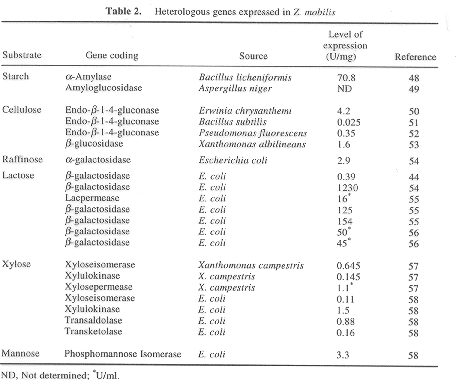
Manipularea genetica a lui Z. mobilis- prin manipulare genetica o gama de substraturi de la Z. au putut fi extinse la bioprodusi atractivi industriali din agricultura precum zerul, amidonul si celuloza. Aceasta s-a putut realiza prin transferul genelor codificatoare hidrolazelor potrivite. Diferite gene codifficand enzime au nevoie ptr utilizare de o gama larga de surse de carbon transferata la Z. mobilis (Tabelul 2).
Clonarea genelor la Z. mobilis e efectuata de 3 tipuri de vectori: o larga gama de plasmide gazda, vectori naveta si o gama larga de vectori gazfa modificati. Desi tulpinilde de Z. mobilis pot actiona ca si destinatari pentru un numar mare de plasmide gazda precum RP1, RP4, R68,etc vectorii utilizati in mod obisnuit la clonare vectorii phage nu pot fi transferati sau mentinuti in Z. mobilis. Cei mai frecvent utilizati vectori cu numar de copiere mare in clonare deriva din RSF1010 si s-a observat ca sunt mai stabili in Z. mobilis decat in derivatii plasmidici RP4. Aparitia plasmidelor naturale in Z. mobilis a fost descrisa si s-a gasit un grad ridicat de omologie intre ele. Plasmidele naturale cu masa molara mica prezente in Z. mobilis sunt folosite la dezvoltarea clonarii vehiculelor ptr. Z. mobilis. Ele au fost unite cu plasmide mici de la E. coli, precum pACYC184 si pBR325 , pentru a produce noi vectori naveta. Segmentele bine definite de la E. coli au facilitat intretinerea plasmidei suplimentare, in timp ce portiuni din Z. mobilis wich includes presently undefined sequences yelds stability in Z. m. via the origin of replication or native plasmid. Vectorul plasmid naveta integrativ, pZMOCP1, a fost construit prin legarea EcoRV digests de clona vetorului plasmid, pZMP1, cu EcoRV digested plasmid, pBluescript DNA.
Transferul plasmidului R la Z. mobilis fie de la E.coli, fie de la Pseudomonas este de obicei realizata prin conjugare. Rate ridicate de conjugare au fost raportate pentru transferul plasmidului in tulpini mutante antibiotic-sezitive CP4.45. Conjugarea eficienta este realizata fie de o mobilizare de sine sau plasmid ajutator, ca si RP4,pRK2013, etc.astfel incat sa aiba loc transformarea sistemului lui Z. mobilis in stadiul sau rudimentar. Transferul plasmidului mediat de mobilizarea donorului celulei cromosomale , ca de ex. E.coli S17,1 este foarte ineficienta. Pentru am imbunatati hidroliza zaharozei, genele zaharazei extracelulare, SacB si SacC, au subclonate in E. coli si Z. mobilis vectorul naveta pZA22. Acest segement a fost transferat la Z. mobilis prin conjugare folosind plasmidul ajutator pRK2013. O metoda alternativa pentru transferul genelor in Z. mobilis este prin fuziunea sferoplastului. Yanase si colaboratorii au izolat FUSANTS pe un mediu rafinos prin trecerea fructose-assimilation negative strains, dar fuzantii nu au putut asimila fructoza.
Un operator la lactozei a fost introdus in Z. mobilis, folosind plasmidul pGC91.14 cu transpozomul Tn951 ce codifica operonul. Z. mobilis care contine operonul lactoza, Tn951, produce 0,4% etanoldin 4% lactoza in 40 de h. De cand Z. mobilis poate metaboliza doar portia de glucoza a loctozei si, de asemeanea, sa genereze doar un mol de ATP per molecula de glucoza metabolizata, nu mai exista castig net de energie pentru cresterea celulei asa cum ATP este cheltuit pentru fiecare molecula de lactoza transportata dea lungul membranei prin transport simport. Pentru a rezolva aceasta problema, Buchhol si colaboratorii au sugerat transferul genei b-galactozidaza intr-o tulpina indiscreta de Z. mobilis impreuna cu operonul galactoza, asa incat genele codificatoare pentru captarea si utilizarea completa a lactozei sa fie disponibile.
Un plasmid recombinant gal+, pZG13, a fost construit prin insertia genei galETK de la E. coli, in aval de promotorul pZA22 de la Z. mobilis si a fost introdus intr-o tulpina de Z. mobilis IFO133756. Z. mobilis recombinat a putut prelua galacoza si sa produca o cantitate redusa de etanol.
O gena a amilazei de la Bacillus licheniformis a fos subclonat in pKT210 si plasmidul recombinat pGNB6 a fost transferat la Z. mobilis. Aici, enzima a fost eliberata prin secretie si, duoa aceea, prin liza celulara. Este, de asemenea, necesar clonarea si transferul unei aminoglucozidaze astfel incat ulpina sa poata creste direct pe amidon. Clonarea genei glucoamilazei de la Aspergillus niger a fost incercatam dar transconjugatii stabili nu au putut fi obtinuti la Z. mobilis.
Pentru a converti celuloza direct la etanol, genele celulazei au fost transferate la Z. mobilis. O CMC-aza de la Cellulomonas uda, CB4, a fost clonata pe pZA22 , un vector de clonare ptr. Z. mobilis. Z. mobilis care poseda aceasta gena sintetizeaza celulaze identice imunologic cu cele ale C. uda. Endoglucanazele de la B.subtilis si P. fluorescens au fost subclonate si introduse in Z, mobilis. In toate aceste cazuri, activitatea endoglucanazelor nu a fost detectata in culturile supernatante, si s-a obtinut o expresie slaba in Z. mobilis fata de E.coli. Oricum, celulaza de la E. chrysanthemi codificand endoglucanaza a fost subclonata intr-o gama larga de vector gazda pGSS33, si prin conjugare a fost transferat la Z. mobilis cu ajutorul RP4. In acest caz, cele mai multe endoglucanaze acumulte in spatiul periplasmic, sugereaza un export eficient a cestor proteine straine in Z. mobilis. Activitatea Cm-celulazei a fost legata de celule in timpul fazei de crestere exponentiala, iar dupa 28 de h peste 40% din activitate s-a recuperat din mediul de cultura. O CM celulaza a Acetobacter xylinum a fost subclonata la vectorul naveta plasmid pZA22 Z. mobilis - E. coli. Plasmidul recombinat rezultat, pZAAC21 a fost introdus intr-un ORF 13756 de la Z. mobilis prin electroforeza. Desi exprimarea Cm celulazei in Z. mobilis a crescut de 10 ori, nu a fost reusita nici o incercare de a utiliza aceasta tulpina in producerea etanolului din celuloza.
Transferul b-glucozidazei de la Xanthomonas albilineans la Z. mobilis a fos reusita prin subclonarea b-glucozidazei in pKT404 urmata de imperechere triparentala care implica plasmidul ajutator pRK2013. Cantitatea de b-glucozidaza produsa de tulpinile recombinante ZM6901 si ZM6902 a fost de 7.5, respectiv 10% din cea exprimata in E. coli. ZM 6901 la Z. mobilis a produs 13.3 mM (ml oare?) etanol din 5 mM celobioza dupa 3 zile, utilizand intregile celule.
Swings, J. and De Ley, J., in Bergies Manual of Systematic Bacteriology (eds Krieg, N. R. and Holt, J. S.), William and Wilkins, Baltimore, USA, 1984, vol. 2, pp. 576
Montenecourt, B. S., in Biology of Industrial
Organisms (eds Demain, A. L. and Soloman, N. A.), Benjamin-Cummings
Publishing Co.,
|
Politica de confidentialitate |
| Copyright ©
2026 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |