
Asa cum deja s-a aratat anterior, sistemele celulare au o structura mai complexa in raport cu sistemele necelulare. In cazul produselor de origine vegetala, elasticitatea tesuturilor de legatura si a peretilor celulelor este mai mica, iar continutul de apa este mai mare decat in cazul produselor de origine animala.
In general, procesele de racire pana la temperaturi apropiate punctului de congelare nu au influente asupra tesuturilor vegetale.
In unele cazuri, daca viteza de racire este foarte mare, se poate produce o marire a permeabilitatii membranelor celulare si o scadere a turgescentii. Acest fenomen este denumit soc termic.
In conditii curent intalnite in practica, subracirea produselor se poate produce pana la valori cu cateva grade sub punctul de congelare, dar pentru durate care nu depasesc in general cateva secunde. In cazul in care produsul este acoperit cu parafina, care impiedica procesul de nucleere a ghetii la suprafata, subracirea poate fi mentinuta o perioada lunga de timp la valori de -4°C-5°C sub punctul de congelare.
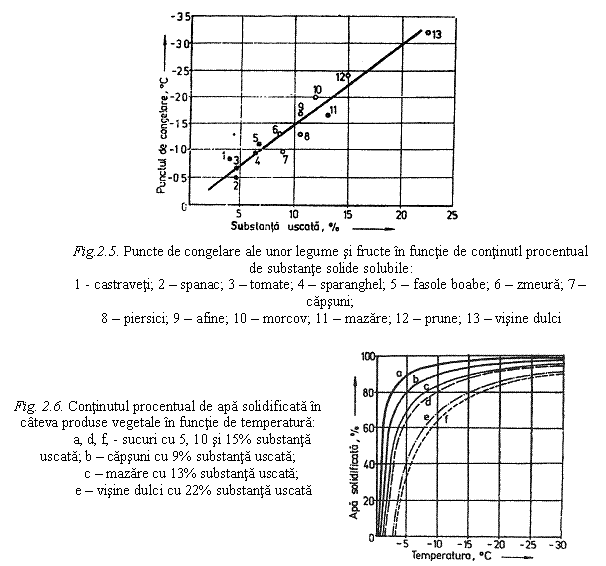
In timpul procesului de congelare, apa exterioara celulelor active din punct de vedere metabolic, congeleaza inaintea apei interioare protoplastelor. Deoarece la aceeasi temperatura apa intercelulara subracita are o presiune de vapori si o energie libera mai mari decat ale ghetii intercelulare, deja formate, apa intracelulara migreaza prin peretii celulelor, producand astfel cresterea cristalelor de gheata intercelulare. Acest proces de migrare a apei continua atat timp cat apa din interiorul celulelor ramane nesolidificata, iar presiunea de vapori este mai mare decat a ghetii exterioare. Transportul apei intracelulare subracite se produce cu atat mai intens cu cat tesuturile vegetale sunt racite mai lent, iar depozitarea acestora se face la temperaturi mai ridicate, sub punctul de congelare. Punctele de congelare ale tesuturilor vegetale comestibile sunt determinate de concentratia in substante solide solubile, in special zaharuri, saruri si acizi. Substantele solide solubile din tesuturile vegetale sunt cuprinse, in general intre 4 si 23%, dintre acestea zaharurile fiind predominante. Pe masura ce temperatura tesuturilor vegetale scade sub punctul de congelare, continutul acestora in gheata creste, iar concentratia in substante solubile si in suspensii ale fazei lichide ramase nesolidificate cresc. Cantitatea de gheata in tesuturile vegetale la o temperatura oarecare, sub punctul de congelare, este determinata de continutul initial de substante solide solubile. In fig. 2.5. este reprezentata dependenta dintre continutul de substante solide solubile si punctul de congelare, iar in fig. 2.6. este redata variatia continutului de apa solidificata in functie de nivelul temperaturii, pentru cateva fructe si legume.
Racirea tesuturilor vegetale pana la temperaturi superioare punctului de congelare nu determina, in general modificari ale structurii si texturii.
In cazul unor durate mai mari ale racirii si in timpul depozitarii in stare refrigerata, pot aparea insa modificari datorita deshidratarii pronuntate, deshidratare determinata de procesul de transpiratie. Turgescenta naturala initiala a legumelor frunzoase este influentata negativ de pierderile de apa, legumele suferind o vestejire care poate sa conduca la moartea celulelor. Vestejirea accentuata poate determina o inmuiere exagerata si o scadere a consistentei. De asemenea, utilizarea unor temperaturi excesiv de scazute la depozitarea in stare refrigerata a unor varietati de legume si fructe poate conduce la modificari de consistenta, culoare s.a. ca urmare a unor procese biochimice enzimatice.
In cazul congelarii, formarea de cristale de gheata poate determina modificari ireversibile asupra peretilor celulelor si protoplastelor. Ca urmare a acestor modificari, proprietatile de textura ale produselor congelate pot fi net inferioare in raport cu produsele necongelate. Tipurile si intensitatea modificarilor sunt in functie de localizarea si dimensiunile cristalelor de gheata, care, la randul lor, depind de viteza de congelare si conditiile de depozitare a produselor congelate. In cazul unor viteze mici de congelare, cristalele de gheata sunt localizate predominant in zonele extracelulare. Aceste zone contin si vapori de apa care condenseaza pe peretii celulelor in picaturi foarte fine si se solidifica sub forma de cristale microscopice de gheata. In timpul cristalizarii exclusiv extracelulare se formeaza cristale de dimensiuni mari, apa din celule este disclocata si se poate produce spargerea peretilor celulelor, modificand in mod nedorit ultrastructura tesutului. Astfel, in cazul capsunilor, la o congelare lenta in aer, peste 90% dintre celule au pereti distrusi, in timp ce la o congelare rapida, doar 10% din celule au pereti distrusi. Pe masura pierderii din apa intracelulara in timpul congelarii extracelulare, concentratia sarurilor anorganice din protoplaste creste pana la limita de la care incepe precipitarea proteinelor.
In cazul unor viteze relativ mari de congelare, mai mari decat 10°C/min, predomina formarea de cristale intracelulare. In acest caz, atat in spatiile extracelulare cat si in interiorul celulelor se formeaza cristale mici de gheata, spatiile dintre celule nu-si maresc volumul, peretii celulelor nu se distrug, iar protoplasma ramane inchisa intre peretii celulei.
In timpul depozitarii in stare congelata si in prima faza a decongelarii tesuturilor vegetale, se produce o crestere a dimensiunilor cristalelor de gheata. Fenomenul se numeste recristalizare si poate avea un efect negativ, asupra celulelor, conducand la modificari ale structurii si o pierdere a fermitatii tesuturilor.
Proprietatile de textura sunt elemente importante ale calitatilor legumelor si fructelor. Prin textura se intelege un complex de caracteristici: rezistenta la masticare, calitatea suprafetei din punct de vedere al masticarii, fibrozitatea s.a. Aceste caracteristici detectabile senzorial sunt determinate de catre structura celulara si compozitia chimica a tesutului. Ca elemente de comparatie sunt considerate fructele, legumele proaspete, recoltate in stare de maturitate deplina. Proprietatile de textura sunt influentate in mod negativ la viteze mici de congelare si la durate mari de depozitare in stare congelata.
In timpul congelarii si depozitarii in stare congelata, in tesuturile vegetale se produc o serie de reactii chimice care au implicatii asupra calitatii produselor. Cele mai importante dintre acestea sunt: reactii asociate cu modificarile de aroma si gust, si respectiv, reactii asociate cu degradarea pigmentilor, degradarea clorofilei si oxidarea acidului ascorbic.
Modificari de aroma si gust Modificarile de aroma ale legumelor si fructelor sunt determinate in special de procese enzimatice. Astfel de modificari se pot produce in timpul depozitarii legumelor si fructelor care nu au fost supuse unor tratamente de inactivare enzimatica, cum este blansarea sau au fost tratate insuficient. In aceste cazuri produsele metabolice rezultate in urma glicozei (compusi volatili cum ar fi etanolul) se acumuleaza in tesuturi, antrenand modificari de aroma specifica. Modificarile de aroma din timpul depozitarii se resimt si dupa decongelarea si prepararea produselor. Modificari de aroma si gust in legumele si fructele congelate neblansate sau incomplet blansate pot fi cauzate si de oxidarea enzimatica a lipidelor. Un rol important, de catalizator, in oxidarea lipidelor il joaca lipoxigenaza.
Brunificarea enzimatica. In tesuturile vegetale congelate, formarea de pigmenti nedoriti (brunificarea) este determinata de oxidarea enzimatica a compusilor fenolici in prezenta oxigenului. Brunificarea se poate observa la legumele si fructele congelate, depozitate si decongelate cum ar fi: mere, piersici, pere, visine, tomate, ciuperci, sfecla s.a. Brunificarea tesuturilor vegetale congelate este mult mai intensa la suprafata sau in straturile apropiate acesteia datorita prezentei oxigenului atmosferic. In straturile interioare, brunificarea se poate produce in unele produse decongelate datorita prezentei oxigenului in spatiile intercelulare.
Distrugerea peretilor celulelor in timpul congelarii datorita cristalelor de gheata, poate declansa brunificarea enzimatica datorita posibilitatii contactului 0 - difenoloxidazei cu substraturile acesteia. Brunificarea in tesuturile vegetale congelate poate fi redusa sau prevenita prin inactivarea termica a enzimelor, adaugarea de inhibitori de brunificare si eliminarea oxigenului. Deoarece, in cele mai multe cazuri, fructele se consuma nepreparate, evitarea brunificarii se poate realiza prin aplicarea de tratamente termice de inactivare enzimatica. Totusi unele fructe, cum ar fi piersicile, caisele si merele, atunci cand sunt utilizate in unele preparate culinare de tip desert, pot fi blansate, fara a genera prin aceasta modificari inacceptabile.
Ca inhibitori de brunificare se utilizeaza bioxidul de sulf, sulfiti, acid sulfuros, acid citric, acid malic, acid ascorbic s.a.
Degradarea clorofilei. In timpul depozitarii in stare congelata a legumelor verzi care au fost tratate termic inante de congelare pentru inactivare enzimatica, la temperaturi peste -18°C, se produce o modificare lenta a culorii, de la verde stralucitor - imediat dupa congelare - catre verde cenusiu, apoi verde masliniu si in final catre verde maroniu. In cazul legumelor verzi neblansate, viteza de modificare a culorii este mult mai mare.
Desi blansarea este necesara pentru a inhiba degradarea clorofilei in timpul depozitarii in stare congelata, ea nu este o metoda perfecta deoarece chiar in timpul blansarii se produce o oarecare degradare a clorofilei. Intensitatea acestui proces de degradare a clorofilei in timpul blansarii depinde de raportul dintre continutul de clorofila a si clorofila b, durata si temperatura de blansare si valoarea pH-ului.
Oxidarea acidului ascorbic. Acidul ascorbic dizolvat in solutii apoase in care se gaseste dizolvat oxigen, trece prin oxidare in acid dehidroascorbic care, in continuare, prin oxidare, trece ireversibil in acid L-digetogluconic si in produsele sale de degradare.
Viteza de degradare a acidului ascorbic este direct proportionala cu radacina patrata din concentratia in cupru si invers proportionala cu patratul concentratiei de ioni de hidrogen din solutie. Reactia este catalizata de ascorbatoxidaza care se gaseste in mod natural in tesuturile vegetale. Pentru inhibarea reactiei, ascorbatoxidaza trebuie inactivata. Viteza de oxidare a acidului ascorbic continut de legume si fructe depinde de nivelul temperaturii de depozitare si de valoarea pH-ului. Cu cat temperatura de depozitare este mai ridicata si valoarea pH-ului mai coborata, cu atat viteza de oxidare este mai ridicata.
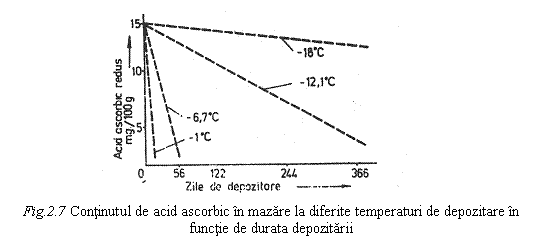
Din fig. 2.7. se observa ca pentru cazul mazarii, cantitatea de acid ascorbic redus, pentru o aceeasi durata de depozitare, este mult mai mare la temperatura de depozitare de -18°C in raport cu temperatura de depozitare de -6°C. Similar este cazul capsunilor care pot fi depozitate cel putin 12 luni la o temperatura de -18 °C fara ca acidul ascorbic sa sufere modificari, in timp ce, depozitate la -12°C timp de 6 luni, jumatate din cantitatea de acid ascorbic se oxideaza.
Modificari ale pH-ului. Asa cum s-a aratat mai inainte, in timpul procesului de congelare a tesuturilor vegetale, se produce o concentrare in substante solide dizolvate in faza lichida intracelulara, necongelata inca. Ca urmare a acestui lucru, unele saruri din compozitia celulelor pot atinge punctul lor eutectic si deci pot sa se separe din solutie conducand la o modificare a pH-ului fazei lichide. Astfel, separarea cifratului de sodiu si de potasiu, conduce la cresterea pH-ului, in timp ce separarea fosfatilor de sodiu conduce la scaderea pH-ului.
Este de subliniat faptul ca modificarile relativ mici ale pH-ului pot determina o alterare a activitatii enzimatice cu efecte negative asupra membranelor celulelor
In timpul racirii si depozitarii legumelor si fructelor, procesele fiziologice continua sa se desfasoare dar cu intensitati mult mai mici in raport cu perioadele anterioare de crestere si maturare. Intensitatea proceselor biochimice care au loc in legume si fructe este determinata de intensitatea procesului de respiratie.
Respiratia consta dintr-un ansamblu de reactii complexe de oxidare a unor substante organice. In cazul respiratiei aerobe oxidarea compusilor organici (in special glucide si acizi organici) are loc pe baza oxigenului atmosferic. In urma procesului de respiratie aeroba rezulta dioxid de carbon si apa, eliminandu-se totodata si o anumita cantitate de energie sub forma de caldura. In functie de natura substantelor oxidate raportul dintre cantitatea de dioxid de carbon degajata si cea de oxigen consumata, denumit si coeficient respirator, are valori diferite. Intensitatea procesului de respiratie poate fi exprimat prin cantitatea de dioxid de carbon degajata din unitatea de masa, in unitatea de timp si este influentata de nivelul temperaturii de pastrare (tabelul 2.1.).
In urma procesului de respiratie se degaja o anumita cantitate de energie sub forma de caldura care este functie de specie, varietate si nivelul temperaturii (tabelul 2.2.).
Pentru asigurarea calitatii produselor depozitate este necesar ca instalatia de racire sau de ventilatie aferenta depozitului de legume si fructe sa fie astfel dimensionata incat sa poata asigura preluarea caldurii degajate in urma procesului de respiratie.
Tabelul 2.1. Influenta nivelului de temperatura asupra intensitatii procesului de respiratie la unele legume si fructe
|
Nr. crt. |
Produsul |
Intensitatea respiratiei, mg CO2/kg h la temperatura de: |
||||
|
0°C |
5°C |
10oC |
15°C |
20°C |
||
|
Ardei | ||||||
|
Bame | ||||||
|
Cartofi | ||||||
|
Castraveti | ||||||
|
Ciuperci | ||||||
|
Ceapa uscata | ||||||
|
Ceapa verde | ||||||
|
Conopida | ||||||
|
Fasole pastai | ||||||
|
Morcovi | ||||||
|
Mazare pastai | ||||||
|
Mazare boabe | ||||||
|
Patrunjel frunze | ||||||
|
Pepeni galbeni | ||||||
|
Ridichi de luna | ||||||
|
Tomate | ||||||
|
Varza alba | ||||||
|
Usturoi |
49,4 | |||||
|
Vinete | ||||||
|
Cirese | ||||||
|
Mere | ||||||
|
Pere | ||||||
|
Prune |
| |||||
|
Piersici | ||||||
Ca urmare a oxidarii substantelor organice, in urma respiratiei, rezulta o anumita cantitate de apa care este functie de natura substantei care se oxideaza. Astfel, in urma oxidarii unei molecule de glucoza rezulta 6 moleculede apa, in urma oxidarii unei molecule de acid malic rezulta 3 molecule de apa, iar oxidarea unei molecule de acid oleic produce 17 molecule de apa.
Apa rezultata din respiratie se poate acumula in celule, poate fi utilizata in procese de sinteza sau se poate elibera impreuna cu apa libera din tesuturi, prin procesul de transpiratie. Procesul de transpiratie este influentat de intensitatea procesului de respiratie, de continutul de apa libera din tesuturi, conditiile de microclimat (viteza, temperatura si umiditatea relativa ale aerului). Cu cat viteza aerului este mai mare, temperatura mai ridicata si umiditatea relativa a aerului mai scazuta, cu atat intensitatea procesului de transpiratie este mai mare.
Respiratia anaeroba sau intramoleculara foloseste, in procesele deoxidare, oxigenul rezultat din degradarea substantelor organice si afecteaza doar glucidele.
Tabelul 2.2. Influenta nivelului de temperatura asupra cantitatii de caldura degajate de legume si fructe prin respiratie
|
Nr. crt. |
Produsul |
Cantitatea de caldura degajata, mg kJ/kg -24h la temperatura de: |
||||
|
0°C |
5°C |
10°C |
15°C |
20°C |
||
|
Ceapa | ||||||
|
Cartofi | ||||||
|
Fasole verde | ||||||
|
Mazare | ||||||
|
Morcovi | ||||||
|
Pepene galben | ||||||
|
Praz | ||||||
|
Ridichi | ||||||
|
Salata | ||||||
|
Spanac | ||||||
|
Sparanghel | ||||||
|
Sfecla rosie | ||||||
|
Telina | ||||||
|
Tomate | ||||||
|
Usturoi | ||||||
|
Varza alba | ||||||
|
Varza rosie | ||||||
|
Agrise | ||||||
|
Banane nematurate | ||||||
|
Caise | ||||||
|
Cirese | ||||||
|
Capsuni | ||||||
|
Mure | ||||||
|
Mere | ||||||
|
Nuci | ||||||
|
Portocale | ||||||
|
Piersici | ||||||
|
Pere | ||||||
|
Prune | ||||||
|
Struguri | ||||||
Intensitatea respiratiei anaerobe este functie de starea de maturare, potentialul oxidoreducator s.a., fiind mai mare pe masura maturarii produselor. Urmare a procesului de respiratie anaeroba, in produse se formeaza si se acumuleaza aldehida acetica si alcool etilic. Daca concentratia de aldehida acetica in tesuturi depaseste 0,004% sau cea de alcool etilic depaseste 0,3% se poate produce moartea celulelor. Respiratia anaeroba este favorizata de reducerea continutului de oxigen din microclimatul in care se gasesc depozitate legumele si fructele. Daca acest continut de oxigen scade sub anumite limite, intensitatea proceselor de respiratie anaeroba devine inacceptabil de mare, conducand la acumulari toxice in tesuturi care pot determina moartea acestora. Respiratia anaeroba se intensifica de asemenea si in cazul unei concentratii prea mari de dioxid de carbon in microclimatul depozitului, a pierderii permeabilitatii la gaz ca rezultat al modificarilor pectinei in timpul maturarii sau dupa maturare si a vatamarii tesuturilor care determina reactii enzimatice anormale.
Maturarea reprezinta un complex de procese fiziologice care conduc in final la atingerea caracteristicilor organoleptice optime si a proprietatilor specifice produsului respectiv care il fac propriu consumului. La marea majoritate a legumelor procesul de maturare continua si dupa recoltare in stransa legatura cu procesul de respiratie si sub influenta directa a conditiilor de microclimat. In functie de stadiul in care se gaseste procesul de maturare in momentul recoltarii si introducerii fructelor si legumelor in spatiul de depozitare precum si in functie de conditiile de microclimat existente in depozit, se pot intalni urmatoarele situatii:
desi temperaturile de refrigerare incetinesc procesele de maturare, in perioada depozitarii se desavarseste maturarea, astfel incat, la scoaterea din depozit, fructele prezinta un maxim de calitate din punct de vedere organoleptic si comercial (in unele cazuri de depozitare a perelor, citricelor s.a.);
la temperaturi sub o anumita limita, unele fructe nu se mai matureaza in timpul depozitarii, astfel incat, dupa scoaterea din depozit, fructele trebuie tinute o perioada de timp la temperaturi mai ridicate inainte de a fi livrate. Astfel de situatii se pot intalni in cazul bananelor, perelor, prunelor, tomatelor s.a.;
in cazul unor legume care in mod intentionat sunt recoltate in faze de crestere, in perioada depozitarii, procesul de maturare nu mai are loc. La scoaterea din depozit in vederea consumului, produsele se gasesc practic in aceeasi faza (de crestere) ca aceea pe care au avut-o la recoltare.
|
Politica de confidentialitate |
| Copyright ©
2025 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |