
DETERMINAREA SPECTRULUI DE SENSIBILITATE LA ANTIBIOTICE A SPECIILOR MICROBIENE. ANTIBIOGRAMA
Datorita specificitatii lor de actiune, antibioticele manifesta eficienta diferita fata de diferite specii microbiene; totalitatea speciilor microbiene sensibile la un anumit antibiotic defineste spectrul de activitate al antibioticului respectiv. In functie de numarul si diversitatea speciilor microbiene afectate, spectrul de activitate al antibioticelor poate fi :
larg (de exemplu, spectrul de actiune al tetraciclinei este reprezentat de bacterii Gram-negative, inclusiv chlamidii si rickettsii si specii Gram-pozitive; penicilinele sunt active in special fata de specii Gram-pozitive, dar si Gram-negative, inclusiv chlamidii; nitrofuranii, rifampicina, sulfamidele sunt active pe un numar mare de specii bacteriene Gram-pozitive si Gram-negative si pe bacteriile acido-alcoolo-rezistente);
ingust (novobiocina este activa pe bacteriile Gram-pozitive, mai ales stafilococi, dar si pe coci si bacili Gram-negativi, cum ar fi hemofilii si pasteurelele; glicopeptidele, bacitracina, pe bacterii Gram pozitive);
limitat (nitroimidazolii sunt activi doar pe microorganismele anaerobe).
Chiar in cadrul aceleiasi specii microbiene, pot exista diferente mari de sensibilitate a diferitelor tulpini fata de un anumit antibiotic, astfel ca stabilirea tratamentului cu antibiotice in clinica necesita izolarea tulpinii microbiene care reprezinta agentul etiologic al infectiei respective (mai ales daca acesta apartine unor genuri si specii supuse fenomenului de dobandire a rezistentei clinice) si determinarea spectrului sau de sensibilitate la antibiotice.
Evaluarea in vitro a eficientei unui antibiotic se realizeaza prin masurarea a 3 parametri, care variaza in functie de concentratia antibioticului si de timpul sau de actiune:
concentratia minima activa (= C.M.A.) - este concentratia la care antibioticul poate induce anumite perturbari in activitatea metabolica a microorganismelor, fara a afecta capacitatea de multiplicare si viabilitatea microorganismelor;
concentratie minima inhibitorie (= C.M.I.) - este concentratia la care un antibiotic inhiba multiplicarea microorganismelor (efect bacteriostatic);
concentratie minima bactericida (= C.M.B.) - este concentratia la care un antibiotic are actiune letala asupra microorganismelor (efect bactericid).
Cunoasterea acestor parametri prezinta o importanta clinica deosebita, deoarece, pentru unele antibiotice, concentratia bactericida este foarte greu de atins in vivo. Din acest motiv, unele antibiotice ca tetraciclina, cloramfenicolul, rifampicina sunt considerate antibiotice bacteriostatice.
Pe langa valoarea C.M.I. determinata in vitro, in alegerea tratamentului cu un anumit antibiotic, trebuie sa se tina cont si de calitatile farmacologice ale antibioticului respectiv (de exemplu, posibilitatea realizarii concentratiei active la nivelul focarului de infectie), de eventualele efecte secundare si de starea fiziologica a bolnavului.
Tehnica de evidentiere a sensibilitatii la antibiotice a unei tulpini microbiene se numeste antibiograma. Antibiograma trebuie practicata in mod obligatoriu in cazul microorganismelor patogene, supuse fenomenului de dobandire a rezistentei, inaintea inceperii oricarui tratament cu antibiotice.
Dupa determinarea spectrului de sensibilitate la antibiotice prin tehnici calitative (in care tulpina microbiana de testat este pusa in contact cu diferite antibiotice aflate intr-o anumita concentratie), se pot practica teste cantitative pentru determinarea valorii C.M.I. (in care tulpina microbiana este pusa in contact cu concentratii crescatoare ale aceluiasi antibiotic).
Metoda difuzimetrica (Kirby-Bauer)
Este o metoda foarte simpla si rapida, care permite determinarea concomitenta a spectrului de sensibilitate a microorganismului si a valorii C.M.I. in vederea calcularii dozelor terapeutice de antibiotice. Metoda are mai multe variante, in practica folosindu-se curent tehnica discurilor impregnate cu antibiotice, standardizata, recomandata de NCCLS (National Committee for Clinical Laboratory Standardisation).
O serie de factori, ca de exemplu, tulpina microbiana studiata (densitatea inoculului, specia, varsta culturii), mediul de cultura (compozitia mediului, pH-ul, densitatea si grosimea stratului de mediu), tehnica folosita si criteriile de interpretare a rezultatelor obtinute, pot influenta rezultatele unei antibiograme. Din acest motiv, tehnica trebuie efectuata in conditii standardizate, reproductibile, conform indicatiilor forurilor internationale in domeniu.
Pe suprafata unui mediu agarizat insamantat in panza cu un inocul standardizat, obtinut din tulpina de testat, se plaseaza la distante egale discuri impregnate cu solutii de antibiotice de o anumita concentratie care vor difuza in mediu, realizand un gradient de concentratie invers proportional cu diametrul zonei de difuzie, deci cu distanta fata de disc. Daca tulpina este sensibila la un anumit antibiotic, cresterea microbiana va fi inhibata pe o anumita suprafata in jurul discului impregnat cu antibioticul respectiv, suprafata denumita zona de inhibitie a cresterii.
Citirea rezultatelor se realizeaza prin masurarea diametrelor zonelor de inhibitie a cresterii determinate de diferite antibiotice, cu ajutorul unei rigle gradate. In cazuri de urgenta clinica se poate realiza o prima citire la 6-8 h de la incubare. Interpretarea rezultatelor se face in functie de dimensiunea zonelor de inhibitie a cresterii, exprimand rezultatul cu termenii de tulpina sensibila (S), rezistenta (R) sau intermediar sensibila (I), conform tabelelor cu puncte critice standardizate si corespunzatoare metodei de lucru: tabelele NCCLS pentru metoda difuzimetrica recomandata de NCCLS (National Committee for Clinical Laboratory Standards, USA), in prezent CLSI (Clinical laboratory and Standards Institute)
Termenii S, I, R definesc de fapt si categoriile de antibiotice, in functie de efectul lor clinic, dupa cum urmeaza:
v categoria S, inseamna ca exista o mare probabilitate ca antibioticul, administrat in doze obisnuite, sa elimine infectia determinata de tulpina testata (C.M.I. are valori net inferioare celor ale concentratiilor umorale obtinute in urma administrarii unei doze obisnuite);
v categoria I, semnifica probabilitatea ca antibioticul sa fie eficient in vivo prin administrare locala sau prin realizarea in mod fiziologic a concentratiilor mari in organe sau tesuturi (rinichi, ficat, cai biliare), la nivelul carora este localizat procesul infectios;
v categoria R inseamna ca, cel mai probabil, administrarea antibioticului nu va determina eliminarea din organism a agentului infectios, a carui sensibilitate a fost testata sau rezultatul tratamentului este imprevizibil.
La citirea si interpretarea rezultatelor se iau in considerare diametrele zonelor de inhibitie, lipsite complet de colonii vizibile cu ochiul liber.
Aparitia coloniilor la marginea sau in interiorul zonei de inhibitie se poate datora urmatorilor factori: cultura este mixta sau suprainfectata; cultura este pura, dar prezinta celule heterorezistente; aparitia mutantelor rezistente; dezvoltarea tardiva a unor celule, de fapt sensibile, si aparitia coloniilor dupa ce antibioticul s-a diluat prin difuzie in mediu.
2. Metode cantitative de determinare a valorii C.M.I.
2.1. Metoda dilutiilor in mediul lichid sau in agar
Principiul metodei: un inocul standardizat al tulpinii testate este insamantat intr-un gradient discontinuu de concentratii ale antibioticului, fie in placi cu mediu agarizat, fie in tuburi cu bulion nutritiv. Dupa incubarea adecvata, se citeste valoarea C.M.I. prin observarea macroscopica a tuburilor: in primele tuburi, cu concentratii mari de antibiotic, cresterea culturii nu este vizibila, microorganismele fiind omorate sau inhibate in prezenta antibioticului. Concentratia de antibiotic corespunzatoare tubului cu cea mai mica concentratie, care inhiba cresterea vizibila a culturii microbiene, reprezinta valorea C.M.I. (mcg/ml) pentru antibioticul respectiv. In tuburile urmatoare, inclusiv tubul martor de crestere, mediul se tulbura ca urmare a cresterii microbiene. In tubul martor de sterilitate, obligatoriu mediul trebuie sa ramana steril (limpede), iar pe placile cu mediu agarizat, coloniile lipsesc. Determinarea C.M.I. se utilizeza pentru stabilirea dozei terapeutice si a caii de administrare in cazul infectiilor severe, supravegherea evolutiei rezistentei bacteriilor la antibiotice, cuantificarea activitatii bactericide a substantelor antimicrobiene. Aceasta metoda permite si aflarea valorii C.M.B. (concentratia minima bactericida) pentru antibioticul testat. Pentru aceasta, se preleveaza 0.01 ml sau 0.1 ml din tuburile utilizate pentru tehnica dilutiilor in mediu lichid (din tubul la care s-a stabilit valoarea C.M.I. si din tuburile anterioare care prezinta concentratii superioare de antibiotic) si se insamanteaza pe suprafata unor placi cu mediu solid nesuplimentat cu antibioticul testat. Dupa incubare, se va observa dezvoltarea microorganismelor la dilutia corespunzatoare C.M.I. Valoarea C.M.B. este data de cea mai mica concentratie de antibiotic care reduce numarul coloniilor pana la 99.9%.
2.2. Metoda E-test
La ora actuala, exista variante ale metodei difuzimetrice pentru determinarea valorii C.M.I.. O astfel de varianta este E-testul (Epsilometer test), ce utilizeaza benzi impregnate cu diferite antibiotice ale caror concentratii variaza exponential si sunt inscrise pe banda respectiva. Zona de inhibitie a cresterii microorganismului testat are aspect de elipsa, al carei diametru variaza direct proportional cu gradientul de concentratie a antibioticului difuzat in mediu, diminuandu-se odata cu scaderea concentratiei astfel ca, la o anumita valoare, zona de inhibitie a cresterii va intersecta banda, concentratia inscrisa pe banda la acest nivel indicand valorea C.M.I.
3. Controlul de calitate in testarea sesibiltatii la antibiotice
Obiective:
- QC (quality control)are drept scop sa asigure:
- precizia si fiabilitatea tehnicii de testare a sensibilitatii
- performanta reactivilor utilizati
- performanta personalului care efectueaza testele.
Pentru a raspunde acestor deziderate, laboratoarele care realizeaza antibiograma trebuie sa dispuna de tulpini de referinta :
- Escherichia coli ATCC 25922 CIP 7624
- Escherichia coli ATCC 35219
- Staphylococcus aureus ATCC 25923 CIP 7625
- Staphylococcus aureus ATCC 43300
- Staphylococcus aureus ATCC 25923 CIP 76110
- Enterococcus faecalis ATCC 29212 CIP 103214
- Haemophilus influenzae ATCC 49247
- Haemophilus influenzae ATCC 49766
- Neisseria gonorrhoae ATTC 49226
- Neisseria gonorrhoae ATTC 49226
- Streptococcus pneumoniae ATCC 49619
Proceduri de control
o - Escherichia coli ATCC 25922
o - Staphylococcus aureus ATCC 25923
o - Staphylococcus aureus ATCC 25923
1. citirea si interpretarea diametrelor zonelor de inhibitie
2. mediul de cultura
3. inoculul
4. discurile de antibiotice
5. tulpinile de referinta
- citirea antibiogramei trebuie realizata cu ajutorul unei rigle precise si sa se evite la maximum erorile, mentinandu-se instrumentul de masura perpendicular pe axa optica;
- trebuie verificat daca interpretarile (S, I, R) corespund diametrelor masurate;
- trebuie evitate erorile prin confuzia diferitelor tabele de citire.
Controlul mediului:
pH:
- trebuie sa fie 7,2 - 7,4
- variatiile de pH influenteaza activitatea aminozidelor, macrolidelor si fenicolilor
- controlul trebuie facut pentru fiecare sarja de mediu Mueller Hinton, cu ajutorul unui pH-metru: se plonjeaza electrodul intr-un flacon de MH semilichid (atentie: temperatura nu trebuie sa fie prea ridicata pentru a evita riscul aprinderii electrodului), geloza trebuie sa se solidifice in jurul extremitatii electrodului; in acest moment se inregistreaza pH-ul.
Umiditatea:
- placile cu mediu trebuie sa fie uscate iinainte de inoculare
Concentratia in timidina sau timina:
- o concentratie prea crescuta de timidina determina o reducere a diametrelor zonelor de inhibitie in jurul discurilor de sulfamide si trimetoprim
- pentru aceasta trebuie testat mediul MH cu o tulpina de referinta Enterococcus faecalis ATCC 29212; diametrul de inhibitie in jurul discului de cotrimoxazol trebuie sa fie >/= 20 mm.
Concentratia in cationi divalenti:
- o concentratie prea crescuta in ioni bivalenti (in principal de Ca2+ si Mg2+) determina o reducere a diametrelor zonelor de inhibitie in jurul discurilor de aminozide (testate pentru P. aeruginosa), in timp ce concentratii mai mici favorizeaza aparitia unor zone de inhibitie prea mari
- ionii de Zn influenteaza activitatea carbapenemelor
- concentratiile trebuie sa fie : Ca2+ = 50 100 mg si Mg2+ = 20 - 35 mg.
Controlul inoculului:
- etalonul 0,5 Mc Farland - se poate obtine astfel:
0,5ml Mg Cl2 sol 1% (10 g/l) +
95,5 ml H2SO4 1% (10 ml/ l);
etalonul astfel preparat trebuie sa prezinte o DO de 0,08 - 0,1 la 625 nm.
- se repartizeaza aceasta solutie in volume de 10 ml, in tuburi identice cu acelea care vor servi la prepararea inoculului
- tuburile se inchid ermetic pentru a evita evaporarea (parafilm, adeziv etc.)
- se marcheaza nivelul lichidului cu un marker si se controleaza periodic masurandu-i DO
- se conserva tuburile la temperatura ambianta, ferit de lumina (hartie de aluminiu)
- tubul etalon se agita inainte de a-l compara cu inoculul preparat: inoculul si etalonul trebuie sa aiba aceeasi turbiditate atunci cand sunt comparate pe un fond cu dungi.
Discurile de antibiotice (tabelul 10, 11):
inainte de utilizare, cartusul de antibiotic trebuie verificat pentru data valabilitatii, mai ales pentru b-lactamine, si pentru incarcatura discului;
stocul de cartuse de antibiotice trebuie sa fie pastrat la -20 oC; aplicatorul incarcat cu antibiotice trebuie pastrat la + 4 oC;
discurile umede sau in contact direct cu gheata, sau care au fost conservate la temperatura ambianta nu trebuie utilizate;
cartusele trebuie scoase din frigider sau congelator cu 1-2 h inainte de utilizare.
Tulpinile de referinta:
- de la primirea lor, tulpinile de referinta vor fi izolate pe un mediu adecvat; plecand de la aceasta cultura se fac 12 subculturi care se repartizeaza in 12 tuburi si se conserva prin congelare la - 70 C.
|
Tulpina de referinta |
Mod de conservare |
Observatii |
|
E. coli ATCC 25922, S. aureus ATCC 25923, P. aeruginosa ATCC 27853 |
- liofilizare - congelare la - 70 oC - geloza profunda |
- conservare pe termen lung - conservare pe termen lung - trecere regulata a tulpinii |
|
S. pneumoniae ATCC 49619 |
- liofilizare - congelare la -70 oC |
Alternativ se poate insamanta o GSC* inclinata, incubata 24 h, apoi conservata la 20 oC. Aceasta tehnica permite conservarea pana la 6 luni. |
*GSC = geloza sange
In fiecare luna se va scoate un tub congelat cu tulpina / tulpinile care vor fi testate; se refac 12 tuburi cu culturi pentru congelare, pentru anul viitor, pornind de la al 12-lea tub consevat.
Tabelul 10: Antibiotice de testat pentru cocii Gram-pozitivi
|
Staphylococcus aureus Lista standard Penicilina* Eritromicina Lincomicina Pristinamicina Kanamicina Tobramicina gentamicina cloramfenicol tetraciclina cotrimoxazol acid fusidic rifampicina *- se testeaza pe geloza Mueller Hinton cu 4% NaCl si mg/ml oxacilina |
Streptococcus spp (altii decat S. pneumoniae) Lista standard penicilina G ampicilina sau amoxicilina oxacilina tetraciclina eritromicina lincomicina sau clindamicina pristinamicna clindamicina Antibiotice optionale rifampicina cefotaxim cefepim streptomicina kanamicina gentamicina cloramfenicol spiramicina telitromicina linezolid cotrimoxazol* ofloxacin vancomicina teicoplamina * - se testeaza pe geloza Mueller Hinton cu 5% sange de cal hemolizat |
Streptococcus pneumoniaeLista standard penicilina G ampicilina sau amoxicilina oxacilina cefotaxim sau ceftriaxon tetraciclina eritromicina lincomicina sau clindamicina pristinamicina Antibiotice optionale imipenem) streptomicina kanamicina gentamicina cloramfenicol telitromicina linezolid cotrimoxazol* fluorochinolone fosfomicina vancomicina teicoplamina * - se testeaza pe geloza Mueller Hinton cu 5% sange de cal hemolizat |
Enterococcus spp Lista standard - ampicilina - gentamicina - nitrofurani Antibiotice optionale - oxacilina* - streptomicina - kaanamicina - cloramfenicol - tetraciclina - eritromicina - lincomicina* sau clindamicina* - pristinamicina (E. faecium) - linezolid - cotrimoxazol - rifampicina - fluorochinolone - vancomicina (Hteicoplamina * - ajuta la identificarea E. faecalis (rezistenta naturala) |
Fenotipuri de rezistenta la antibiotice la cocii Gram-pozitivi
|
Streptococi Rezistenta naturala: - nivel scazut la aminozide; polimixine, acid fusidic, acid nalidixic; sensibilitate moderata la fluoroquinolone; sensibilitate scazuta la acid fusidic. NB: Activitatea SXT inhibitata de prezenta timidinei in mediu (geloza Mueller Hinton cu adaos de 5% sange de cal). |
Pneumococi Rezistenta naturala de nivel scazut la aminozide, acid fusidic, polimixina, acid nalidixic, - putin sensibil la pefloxacina si norfloxacina (sensibil la sparfloxacina), bacitracina |
|
Rezistenta dobandita la stafilococi: rezistenta la penicilina G; fenotip MLSb inductibil la eritromicina; fenotip KT. Rezistenta dobandita la streptococi : la toate clasele de antibiotice cu exceptia glicopeptidelor. Rezistenta la penicilina decelata cu ajutorului discului de Oxacilina (5/ 1 µg), incrucisata cu toate beta-lactaminele, dar de niveluri diferite (se calculeaza C.M.I.). |
|
Tabelul 11: Antibiotice de testat pentru Enterobacteriaceae si bacili Gram negativi nonfermentativi
|
Escherichia coli |
Salmonella Amoxicilina/ Ampicilina Gentamicina/ Tobramicina ftazidim/ Cefotaxim/ Ceftriaxon Ofloxacin/ Ciprofloxacin Cotrimoxazol Acid nalidixic |
Klebsiella Amoxicilina/ Ampicilina Geentamicina/ Tobramicina Ceftazidim/ Cefotaxim/ Ceftriaxon Ofloxacin/ Ciprofloxacin Cotrimoxaszol |
P. aeruginosa Acinetobacter spp. Ticarcilina Piperacilina Ceftazidim Imipenem Amikacina Gentamicina Tobramicina Ofoxacin |
Vibrio Ampicilina Cotrimoxazol Acid nalidixic Tetraciclina |
Haemophilus influenzae** Ampicilina/ Amoxicilina Cefalotin Gentamicina Cloramfenicol Pristinamicina Eritromicina Tetraciclina Pefloxacina Sulfametoxazol Rifampicina. |
* inocul McFarland 0,5.in bulion Mueller Hinton sau in solutie salina ( 0,9% Na Cl) pornind de la o cultura de 18 - 24 h pe mediu solid neselectiv
** Rezistenta constitutiva: la aminopeniciline, carboxipeniciline, ureidopeniciline- b-lactamaza sensibila la inhibitori, frecvent asociata cu multirezistenta; la aminopeniciline, cefalosporine generatia I nonenzimatica, sensibilitatea nu este restabilita in prezenta inhibitorilor de beta-lactamaze.
Asadar, antibiograma difuzimetrica poate permite determinarea pattern-urilor de rezistenta constitutiva, intrinseca, nativa, precum si detectarea fenotipica a unor mecanisme de rezistenta dobandita.
Plasarea antibioticelor pe placa intr-o anumita ordine permite citirea interpretativa a testelor de sensibilitate la antibiotice (concept introdus de Patrick Courvalin in 1992) si detectarea fenotipica a unor mecanisme de rezistenta la antibiotice.
Rezistenta S. aureus la β-lactamice
S-au evidentiat mai multe mecanisme de rezistenta la β-lactamice (tabelul 12, 13).
Tabelul 12: Mecanismele de rezistenta la agenti antimicrobieni la S. aureus (dupa Lowy, 2003).
|
Antibiotice |
Genele rezistente |
Produsele genelor |
Mecanismele de rezistenta |
|
ß-lactame |
blaZ mecA |
ß-lactamaza PBA2a |
1) Hidroliza enzimatica a nucleului ß-lactamic (fig. 24) 2) Reduce afinitatea pentru PBP |
|
Glicopeptide |
VISA - |
1) Peptidoglican alterat 2) D-Ala-D-Lac |
1) Legarea vancomicinei in peretele celular 2) Sinteza dipeptidelor cu reducerea afinitatii pentru vancomicina |
|
Quinolone |
1) parC 2) gyrA sau gyrB |
1) Componentul par C al topoizomerazei IV 2) Componentele Gyr A sau GyrB ale girazei |
1, 2) Mutatii care au loc in regiunea QRDR reduc afinitatea complexului ADN-enzima pentru quinolone |
|
Trimethoprim-sulfamethoxazol |
1) Sulfonamide: sal A 2)TMP:dfrB |
1) Dhidropteroat sintaza 2) Dhidrofolat reductaza |
1) Supraproductia de acid amino-benzoic de catre enzime 2) Reducerea afinitatii pentru DHFR |
|
Quinopristin-dalfopristin (Q-D) |
1)Q:ermA ermB ermC 2)D:vat, vatB |
1) Metilaze ribosomale 2) Acetiltransferaze |
1) Reduce legarea la subunitatea ribosomala 23S 2) Modificarea enzimatica a dalfopristinului |
Tabelul 13: Mecanismele de rezistenta la -lactamice la S. aureus (dupa Lowy, 2003).
|
Mecanism |
Penicilina G Penicilina A Carboxipenicilina Ureidopenicilina |
Antibiotic + inhibitor de β-lactamaze |
Penicilina M |
Cefalosporine Carbapeneme |
|
Salbatic |
S |
S |
S |
S |
|
Penicilinaza |
R |
S |
S |
S |
|
Modificarea PLP, gena mecA |
R |
R |
R |
R |
|
BORSA |
R |
S/R |
R |
S |
|
MODSA |
S |
S |
R |
S |
BORSA = S. aureus borderline; MODSA = S. aureus modificat.
Rezistenta la meticilina (respectiv oxacilina) este considerata un marker al polirezistentei incluzand: cefalosporinele, eritromicina si clindamicina, iar izbucnirile intraspitalicesti cu acest tip de tulpini au creat probleme deosebite terapeutice, antiepidemice si de cost al spitalizarii (Codita, 1993).
Detectarea producerii de β-lactamaza este obligatorie pentru toate tulpinile de stafilococ care prezinta un diametru la penicilina mai mare de 28 mm sau tulpinile de N. gonorhoeae sau Hemophilus spp.
- exista mai multe tehnici: acidimetrice, cromogenice si iodometrice
- testele pentru detectarea β-lactamazei trebuie sa fie rapide (implicatii terapeutice) si sa preceada realizarea antibiogramei.
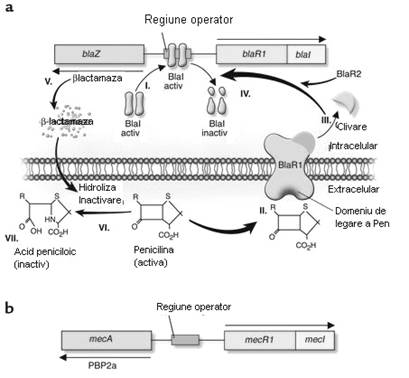
Fig. 24. Inducerea sintezei ß-lactamazei stafilococice in prezenta penicilinei.
BlaI se leaga de regiunea operator, astfel incat se represeaza transcrierea ARN atat pentru blaZ, cat si pentru blaR1-blaI; II Legarea penicilinei la BlaR1 stimuleaza activarea autocatalitica a BlaR1; III-IV BlaR1 cliveaza BlaI in fragmente inactive permitand transcrierea blaZ si blaR1-BlaI; V-VIII ß-lactamaza, enzima extracelulara codificata de blaZ (V) hidrolizeaza ciclul ß-lactamic al penicilinei (VI), in consecinta produce inactivarea (VII); Schema operonului care codifica rezistenta la meticilina a S. aureus. Expunerea MecR1 la un antibiotic ß-lactamic induce sinteza MecR1; MecR1 inactiveaza MecI, permitand sinteza lui PBP2a (dupa Kernodle, 2000).
Testul treflei:
Se insamanteaza o tulpina de S.aureus ATCC 25923 pe o geloza Muller-Hinton (sau M-H cu sange Chocolat pentru N.gonorroeae si Haemophilus); Se aplica un disc de peniciclina G in centrul placii (sau de ampicilina in cazul Haemophilus); Se insamanteaza in striuri radiale (din centrul placii spre periferie) tulpina de testat, o tulpina S. aureus ATCC 25923 (sensibila la penicilina G), o tulpina de S. aureus ATCC 43300 (rezistenta la penicilina G) (fig. 25). Se incubeaza placa 18 h la 35°C in atmosfera normala (sau 24 h in atmosfera imbogatita cu CO2 pentru Haemophilus si N. gonorrhoeae).
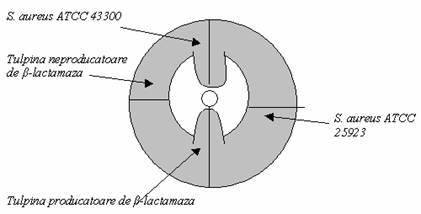
Fig. 25. Reprezentarea testului de detectare a producerii de β-lactamaza prin metoda treflei
(dupa Codita si colab., 2005)
Se suspecteaza existenta β- lactamazei daca se observa o diferenta intre diametrul de inhibitie realizat in cele 2 cazuri, in favoarea AMC, cele 2 diametre fiind in categoria sensibila a valorilor critice admise pentru tulpina testata.
Rezistenta la meticilina in cazul stafilococului auriu este de multe ori asociata cu mecanisme de rezistenta care determina inactivarea altor familii de antibiotice. S. aureus rezistent la meticilina prin modificarea PLP este adesea rezistent la aminozide (amikacina si tobramicina si in peste peste 90% din cazuri la gentamicina), la fluoroquinolone, macrolide, lincosamide si ketolide, la fosfomicina si uneori la rifampicina. Ca regula generala, raman active pe stafilococ glicopeptidele, acidul fusidic, streptograminele, rifampicina si oxazolidinonele.
La enterococi, rezistenta la β-lactamine se realizeaza prin producerea β-lactamazei si prin modificarea tintei (ce confera rezistenta naturala la cefalosporine, oxaciline si monobactami si sensibilitate diminuata la peniciline).
Streptococcus pneumoniae prezinta rezistenta dobandita la penicilina prin producerea de b-lactamaze, evidentiata fenotipic prin testarea sensibilitatii la oxacilina, utilizand discuri de oxacilina (Ox , 5µg). Rezistenta la penicilina este incrucisata pentru toate β-lactaminele, dar prezinta diferite niveluri de expresie in functie de antibiotic. Activitatea amoxicilinei si cefalosporinelor din generatia a treia a este mai putin modificata decat cea a penicilinei G.
In ceea ce priveste cefalosporinele din prima si a doua generatie si cele orale, indiferent de generatie, cu exceptia cefpodoximului, ele sunt mai putin active decat penicilina G.
|
β-lactamine active |
Penicilina G |
b-lactamine mai putin active |
|
Imipenem |
Ampicilina |
Piperacilina |
|
Amoxicilina |
Cefpodoxin |
Cefuroxim |
|
Cefotaxim Ceftriaxon |
Cefoperazon Oxacilina Cefixim Cefaclor Cefotetan |
Rezistenta la β-lactamine este asociata in proportie de peste 50% cu rezistenta la alte antibiotice ca: tetraciclinele, macrolidele, cloramfenicolul sau trimetoprim-sulfametoxazolul (fig. 26). Unele esecuri terapeutice ar putea fi date de fenomenul de toleranta la penicilina, datorat modificarilor controlului activitatii autolitice, ducand in vivo la selectarea mutantelor peni-R (raportul CMB/CMI > 32).
Toate tulpinile cu diametrul de inhibitie <26 mm trebuie testate CMI pentru penicilina, amoxicilina, cefotaxim si toate antibioticele utilizate in tratament.

|
AMX amoxicilina |
CXM cefuroxim |
CTX cefotaxim |
MNO minociclina |
|
CPD cefpodoxim |
CEC cefaclor |
SXT trimetroprim-sulfametoxazol |
|
|
OXA oxacilina |
DTM |
CFM cefixime |
RA rifampicina |
|
C cloranfenicol |
KAN kanamicina forte |
CRO cefpiroma |
PT pristinamicina |

|
AMX |
PIP |
CTX |
OXA |
|
GM |
KAN |
GEN |
SXT |
|
RA |
VA |
TEC |
FOS |
Fig. 26. Tulpini de Streptococcus pneumoniae multi-rezistente la antibiotice (original)
Tulpinile de Streptococcus pneumoniae sunt foarte sensibile la optochin, testul la optochin constituind unul dintre testele simple de identificare sau de confirmare a speciei si de diferentiere fata de ceilalti streptococi α hemolitici (testul Lund 1959). La concentratii de 1 / 500.000 1 / 1.000.000, optochinul are actiune bactericida pentru pneumococi, in timp ce pentru ceilalti streptococi este nevoie de doze mult mai mari de 1 / 5.000. Se insamanteaza tulpina de testat pe o placa cu geloza sange continand optochin 0.05% in paralel cu insaamantarea pe o placa de control fara optochin. In jurul pulberii de optochin apare o zona de inhibitie a cresterii, cu un diametru de peste 20 mm (Fig. 27)
Se pare ca doar un numar redus de specii ( streptococi orali; S. oralis, S. sanguis, S. mitis) prezinta sensibilitate diminuata la penicilina G (dupa CLSI).
Dupa sensibilitatea lor naturala la antibioticele β-lactamice, Enterobacteriile sunt clasificate in 4 grupe. Clasificarea are la baza comportamentul fata de urmatoarele antibiotice β-lactamice: aminopeniciline, aminopeniciline + inhibitori ai β-lactamazelor (IBL), carboxipeniciline, ureidopeniciline, cefalosporine din prima generatie (CIG), cefalosporine din a II a generatie (CIIG), cefalosporine din a III a generatie (CIIIG), cefalosporine din a IV a generatie CIVG (cu spectru larg), CIIIG + IBL, carbapeneme, cefamicine.
Grupa I
In aceasta grupa sunt clasate tulpinile de Escherichia coli, Proteus mirabilis, Salmonella ssp. si Shigella spp. (tabelul 14 ). Aceasta grupa prezinta 4 fenotipuri mai frecvente:
Fenotipul 1: tulpini sensibile la toate β-lactamicele (au fost evaluate la aproximativ 60% din frecventa acestei tulpini);
Fenotipul 2: tulpini producatoare de cefalosporinaze cromosomale de nivel scazut (aproximativ 5% din tulpini).
Fenotipul 3: tulpini producatoare de penicilinaze. Aceasta enzima este in general transferabila si poate fi exprimata la nivele mai mult sau mai putin importante (au o frecventa de aproximativ 30% din tulpini).
Fenotipul 4: tulpini producatoare de penicilinaze rezistente la inhibitori.
Grupa II
Aceasta grupa este reprezentata de tulpinile de Klebsiella spp., Citrobacter koseri, C. amalonaticus, E. Hermannii (tabelul 15).
Fenotipul 1: tulpini rezistente la concentratii scazute de amoxicilina si de ticarcilina.
Fenotipul 2: tulpini producatoare de penicilinaze plasmidiale. Nivelul de producere al acestor penicilinaze este variabil, dar in general scazut.
Fenotipul 3: tulpini producatoare de BLSE β-lactamaze de spectru larg. Aceste tulpini sunt rezistente la o concentratie relativ scazuta de cefalosporine de a III a generatie. Pot fi evaluate frecvent la aceste tulpini la aproximativ 15% din toate speciile de Klebsiella.
Grupa III
Este reprezentata de tulpini de Enterobacter spp., Serratia spp., Proteus indol (+) si Morganella, Providencia spp., Citrobacter freundii (tabelul 16).
Fenotipul 1: tulpini cu rezistenta naturala la amoxicilina si la cefalotin, ca urmare a exprimarii de cefalosporinaze cromosomale;
Fenotipul 2: tulpini rezistente la amoxicilina, cefalotina si la ticarcilina, prin producerea unei penicilinaze plasmidiale;
Fenotipul 3: tulpini rezistente la cefalosporinele de generatia a III a;
Fenotipul 4: tulpini de Proteus vulgaris producatoare de cefalosporinaze sensibile la acid clavulanic (cefuroximaze).
Grupa IV
Grupeaza tulpini de Yersinia rezistente la amoxicilina, ticarcilina, cefalotin si cefoxitin, dar sensibile la pipercilina (tabelul 17).
Tabelele urmatoare exprima diferitele fenotipuri de rezistenta (ca si fenotipurile salbatice) stabilind manifestarea fenotipica a productiei de β-lactamaze in fiecare grup. Cateva fenotipuri particulare pot fi datorate altor mecanisme. Producerea unei enzime de catre o bacterie nu raspunde la legea tot sau nimic ci exista niveluri intermediare de exprimare ce pot fi detectate fenotipic prin citire interpretativa foarte utile in intelegerea dinamicii fenomenului de antibiorezistenta.
Tabelul 14: Fenotipuri de rezistenta la antibioticele beta-lactamice la enterobacteriile din grupa I . E.coli, P.mirabilis, Salmonella spp., Shigella spp.
|
Antibiotic |
Fenotip salbatic |
Penicilinaza nivel scazut |
Penicilinaza nivel ridicat |
|
Aminopeniciline |
S |
R |
R |
|
Aminopeniciline + IBL |
S |
S |
I/R |
|
Carboxipeniciline |
S |
R |
R |
|
Ureidopeniciline |
S |
I/R |
I/R |
|
CIG |
S |
I |
I/R |
|
CIIG |
S |
S |
S/R |
|
CIIIG |
S |
S |
S |
|
CIIIG + IBL |
S |
S |
S |
|
Cefamicine |
S |
S |
S |
|
CIVG (1) |
S |
S |
S |
|
Carbapeneme |
S |
S |
S |
R = rezistent, S = sensibil, I = intermediar, (1) cefalosporine cu spectru larg : cefepim si cefpirom
|
Antibiotic |
Cefalosporinaza1 |
TRI 2 |
BLSE3 |
CHN4 |
|
Aminopeniciline |
R |
R |
R |
R |
|
Aminopeniciline + IBL |
R |
R |
R |
R |
|
Carboxipeniciline |
S |
R |
R |
R |
|
Ureidopeniciline |
S |
R |
R |
R |
|
CIG |
R |
S |
R |
R |
|
CIIG |
S |
S |
R |
R |
|
CIIIG |
S |
S |
R |
R |
|
CIIIG |
S |
S |
S |
R |
|
Cefamicine |
S/R |
S |
S |
R |
|
CIVG |
S |
S |
R |
S |
|
Carbapeneme |
S |
S |
S |
S |
E.coli si Shigella, 2-TEM rezistent la inhibitori, 3-β-lactamaza cu spectru extins, 4-cefalosporinaza de nivel ridicat
Tabelul 15: Fenotipuri de rezistenta la antibioticele beta-lactamice la enterobacteriile din grupa a II a : Klebsiella spp.,Citrobacter koseri, C.amalonaticus, E.hermannii
|
Antibiotic |
Fenotip salbatic (1) |
Penicilinaza nivel ridicat |
BLSE |
CHN |
|
Aminopeniciline |
R |
R |
R |
R |
|
Aminopeniciline +IBL |
S |
R |
R |
R |
|
Carboxipeniciline |
R |
R |
R |
R |
|
Ureidopeniciline |
I |
R |
R |
R |
|
CIG |
S |
R |
R |
R |
|
CIIG |
S |
I/R |
R |
R |
|
CIIIG |
S |
S |
R |
R |
|
CIIIG + IBL |
S |
S |
S |
R |
|
Cefamicine |
S |
S |
S |
R |
|
CIVG |
S |
S |
R |
S |
|
Carbapeneme |
S |
S |
S |
S |
(1) penicilinaza de nivel scazut
Tabelul 16: Fenotipuri de rezistenta la antibioticele beta-lactamice la enterobacteriile din grupa a III a : Enterobacter spp., Serratia spp., Providencia spp., Citrobacter freundii, Proteus vulgaris, P. penneri, Morganella. spp.
|
Antibiotic |
Fenotip salbatic (1) |
Penicilinaza de nivel ridicat |
BLSE |
CHN |
|
Aminopeniciline |
R |
R |
R |
R |
|
Aminopeniciline + IBL |
R (2) |
R |
R |
R |
|
Carboxipeniciline |
S |
R |
R |
R |
|
Ureidopeniciline |
S |
R |
R |
R |
|
CIG |
R |
R |
R |
R |
|
CIIG |
S/R |
R |
R |
R |
|
CIIIC |
S |
S |
R |
R |
|
CIIIG + IBL |
S |
S |
S |
R |
|
Cefamicine |
S/R |
S/R |
S/R |
R |
|
CIVG |
S |
S |
R |
S/I |
|
Carbapeneme |
S |
S |
S |
S |
(1) cefalosporinaza inductibila, (2) cefalosporinaza inductibila numai pentru P. vulgaris si P. penneri
Sinergismul dintre augmentin si cefotaxim si/sau aztreonam si/sau cefepim si/sau cefpirom, permite detectia anumitor BLSE. Cefepimul sau cefpiromul sunt utile in detectia unei BLSE in prezenta unei CHN.
Tabelul 17: Fenotipuri de rezistenta la antibioticele b-lactamice a enterobacteriilor din grupa a IV a: Yersinia enterocolitica
|
Antibiotic |
Fenotip salbatic |
BLSE |
|
Aminopenicilina |
R |
R |
|
Aminopeniciline + IBL |
R |
R |
|
Carbeniciline |
R |
R |
|
Ureidopeniciline |
I/R |
R |
|
CIG |
R |
R |
|
CIIG |
S |
R |
|
CIIIG |
S |
R |
|
CIIIG + IBL |
S |
S |
|
Cefamicine |
S |
S |
|
CIVG |
S |
R |
|
Carbapeneme |
S |
S |
Unele tulpini de K.oxytoca, P.vulgaris, P. penneri si Citrobacter koseri sunt hiperproducatoare de <aztreonamaza>, o penicilinaza cromosomala (tabelul 16).
Tabelul 18: Fenotipuri de rezistenta la b-lactamice a tulpinilor producatoare de aztreonamaza:
|
Antibiotic |
Fenotip |
|
Aminopeniciline |
R |
|
Aminopeniciline + IBL |
R |
|
Carboxipeniciline |
R |
|
Ureidopeniciline |
R |
|
CIG |
R |
|
CIIG |
R |
|
Ceftriaxon |
R |
|
Cefotaxim |
S/R |
|
Ceftazidim |
S |
|
AZTREONAM |
R |
|
Cefamicina |
S |
|
Carbapeneme |
S |
Rezistenta la carbapeneme apare prin urmatoarele mecanisme:
impermeabilitate ( P. mirabilis, E.aerogenes)
impermeabilitate si producere de cefalosporinaza de nivel ridicat ( E. cloacae, E. aerogenes, P. rettgeri)
carbapenemaze : de tip penicilinaza, inactiva asupra CIIIG (E. cloacae); de tip metalo-enzima, care hidrolizeaza CIIIG si carbapenemele.
Rezistenta P. aeruginosa la β-lactamice
La P. aeruginosa, rezistenta naturala este indusa prin mecanisme de impermeabilitate, eflux, producerea enzimelor inactivatoare si modificarea tintelor.
Fenotipul salbatic este legat de producerea unei cefalosporinaze inductibile in prezenta aminopenicilinelor si cefalosporinelor din prima si a doua generatie, cuplata cu o impermeabilitate mai mult sau mai putin marcata si de un sistem de eflux constitutiv, care determina aparitia unui fenotip rezistent la aminopeniciline, ca atare sau asociate cu inhibitori ai β-lactamazelor si la cefalosporine de prima si a II a generatie, cu conservarea sensibilitatii la carboxipeniciline, ureidopeniciline, la anumite cefalosporine de generatia a III a, la monobactami si carbapeneme.
Rezistenta dobandita apare in urma manifestarii mecanismelor enzimatice (penicilinaze, cefalosporinaze, BLSE, metalo-enzime) sau neenzimatice (impermeabilitate, eflux si modificarea tintelor) (tabelul 19 ).
Rezistenta dobandita enzimatica este mediata de:
a) penicilinaze plasmidiale transferabile cum ar fi: PSE, TEM, OXA (nivel scazut sau ridicat) care se materializeaza prin rezistenta la carboxipeniciline, ureidopeniciline, cefoperazona si sensibilitate la: ceftazidim, cefepim, imipenem si inhibitori de β-lactamaze.
b) BLSE:
- tip penicilinaza (TEM, SHV, PER, VEB) care dau sensibilitate la imipenem, carboxipeniciline+ IBL si ureidopeniciline+ IBL
- tip oxacilinaza (OXA-2, OXA-10) : sensibilitate la imipenem
c) cefalosporinaze cromosomale derepresate:
- hiperproduse care confera rezistenta la toate β-lactamicele,cu exceptia cefepimului si imipenemului
- hiperproduse cu nivel inalt care confera sensibilitate doar la imipenem
d) carbapenemaze care dau sensibilitate doar la aztreonam
Rezistenta dobandita neenzimatica este indusa prin:
a) trei sisteme de eflux: MexA, MexB si OprM, constitutiv, confera rezistenta naturala la carboxipeniciline si aztreonam; MexC, MexD si OprJ, confera rezistenta la cefepim si cefpirom; MexE, MexF si OprN, cuplata adesea cu D2, confera rezistenta la imipenem.
b) deficit de porine - se refera la pierderea porinelor - care dau rezistenta variabila la carbapeneme (uneori asociata cu o cefalosporinaza derepresata)
c) modificarea LPS-ului duce la permeabilitate diminuata pentru toate β-lactamicele, cu exceptia imipenemului.
d) modificarea PLP-urilor 2 si 4 caredau rezistenta la imipenem si 3 care genereaza rezistenta la toate β-lactamicele, cu exceptia imipenemului
Tabelul 19: Evidentierea fenotipurilor de rezistenta la betalactamice pe baza citirii interpretative la Pseudomonas aeruginosa
|
Antibiotic |
Fenotip salbatic |
Penicillinaza(1) |
CHN(2) |
|
|
Aminopeniciline |
r |
r |
r |
|
|
Aminopeniciline +IBL |
r |
r |
r |
|
|
Carboxipeniciline |
s |
r |
r |
|
|
Ureidopeniciline |
s |
r |
r |
|
|
Cefalosporine 1a generatie |
r |
r |
r |
|
|
Cefalosporine 2a generatie |
r |
r |
r |
|
|
Cefalosporine 3a generatie |
Cefoperazona |
s |
r |
r |
|
Ceftazidim |
s |
s |
r |
|
|
Cefsulodin |
s |
s |
r |
|
|
Cefalosporine de spectru larg |
s |
s |
Cefepim S/R |
|
|
Aztreonam |
s |
s |
R |
|
|
Carbapeneme |
s |
s |
S |
|
(1) Ticarcilina +acid clavulanic: S ; Piperacilina+tazobactam : S
(2) Cefalosporinaza de inalt nivel
(3) IBL : inhibitor de beta-lactamaza
(4) in functie de nivelul de producere
Tabelul 20: Fenotipuri de rezistenta la β-lactamice la Stenotrophomonas maltophilia
Fenotip |
Hiper- sensibil |
Sensibil CAZ-S/CAZ-R |
Imipenem-S |
Altele |
|
Amoxicilina |
R |
R R |
R |
R |
|
Amoxicilina-AC |
R |
R R |
S |
R |
|
Ticarcilina |
S |
R R |
R |
R |
|
Ticarcilina-AC |
S |
S S |
S |
V |
|
Ceftazidim |
S |
S R |
S |
R |
|
Aztreonam |
S |
R R |
R |
R |
|
Imipenem |
R |
R R |
S |
R |
|
Frecventa (%) |
< 1 |
Activitatea si expresia β-lactamazelor
|
L1 (rezistenta la AC) |
+ |
V |
||
|
L2(sensibilitate la AC) |
++ |
V |
S = sensibil; R = rezistent; I = intermediar; V = variabil; AC= acid clavulanic; CAZ = ceftazidim
Rezistenta la β-lactamine la Acinetobacter spp.
Rezistenta naturala se manifesta aproape fara exceptie la aminopeniciline, cefalosporine de prima generatie si de generatia a II a.
Rezistenta dobandita (tabelul 21) este indusa prin mai multe mecanisme :
producerea de β-lactamaze de tipul penicilinazelor plasmidiale TEM (1 si 2), CARB5
oxacilinaze: OXA21
cefalosporinaze cromosomiale ACE 1 ACE 4 care hidrolizeaza toate cefalosporinele
combinarea mecanismelor de tipul: impermeabilitate, modificarea PLP-urilor, producerea de enzime (induc rezistenta la imipenem)
Tabelul 21: Fenotipuri de rezistenta la β-lactamice la Acinetobacter spp
|
Fenotip |
I |
II |
III |
IV |
V |
|
Mecanism |
Penicilinaza |
Cefalosporinaza |
Penicilinaza+ cefalosporinaza |
Acumulare de mecanisme |
|
|
Carboxi si ureidopeniciline |
S |
R |
S |
R |
R |
|
Cefalosporine din gen. a III a |
S |
S |
R |
R |
R |
|
Aztreonam |
S |
S |
R |
R |
R |
|
Carbapeneme |
S |
S |
S |
S |
R |
|
Frecventa |
Rezistenta la β-lactamine la Haemophilus influenzae
Se datoreaza mai multor mecanisme, cel mai frecvent fiind producerea de β-lactamaze, care a aparut la 30% dintre tulpinile izolate din infectii bronho-pulmonare la adulti. Aceste β-lactamaze sunt plasmidiale de tip TEM, mai rar ROB care inactiveaza aminopenicilinele, carboxi- si ureidopenicilinele. In general raman active asociatia amoxicilina-acid clavulanic si cefalosporinele din generatia a III-a. Producerea β-lactamazelor este usor de evidentiat prin teste cromogenice (nitrocefin).
Un alt mecanism de rezistenta destul de frecvent este modificarea tintei. De obicei sunt tulpini necapsulate, rezistenta datorandu-se alterarii uneia sau mai multor PLP-uri si confera rezistenta de nivel scazut la toate antibioticele din aceasta clasa.
Rezistenta la imipenem
Se pare ca s-ar datora diminuarii permeabilitatii membranei externe printr-un mecanism inca neelucidat. Exista in acest sens printre tulpinile sensibile la ampicilina un numar de tulpini rezistente la imipenem.
5. Metode fenotipice calitative si cantitative de evidentiere a beta-lactamazelor
Identificarea definitiva a acesor enzime este posibila numai prin tehnici de secventiere a proteinelor si genelor, tehnici care sunt totusi mult prea costisitoare pentru a fi utilizate in clinica si diagnostic. Totusi, metode mai simple de evidentiere si caracterizare a β lactamazelor sunt disponibile pentru laboratoarele clinice.
Antibiograma difuzimetrica calitativa clasica (metoda Kirby Bauer)
BLSE pot fi sintetizate in cea mai mare parte de catre enterobacterii si sunt reproduse cel mai adesea in antibiograma prin imaginea uni sinergism (dop de sampanie) intre un disc de cefalosporina de generatia a III-a si /sau monobactam si un disc de amoxicilina + acid clavulanic.
Dar aceasta imagine poate fi discreta sau atipica. Secretia de BLSE va fi suspectata mai intai, de micsorarea tuturor diametrelor de inhibitie la toate cefalosporinele de generatia a 3-a : CTX ≤ 27 mm, CAZ ≤ 22mm, CRO ≤ 25mm, ATM ≤ 27mm (tabelul 22). O tulpina de referinta trebuie testata in aceleasi conditii in scopul verificarii validitatii discurilor de antibiotice. Prezenta unei BLSE indica faptul ca tulpina este rezistenta la toate lactaminele cu exceptia imipenemului si cefamicinei (Fig. 28).
|
Antibiotic |
ESBL tip TEM |
ESBL tip SHV |
|
Amoxicilina / Ampicilina |
R |
R |
|
Amoxicilina + acid clavulanic |
S |
S |
|
Piperacilina |
R |
R |
|
Cefoxitina |
S |
S |
|
Cefuroxima |
S |
R |
|
Cefotaxima |
S |
S / I / R |
|
Ceftazidima |
S |
R |
|
Aztreonam |
S |
R |
|
Cefepima |
S |
S / I / R |
|
Imipenem |
S |
S |
Tabelul nr 22: Diferentierea ESBL tip TEM de ESBL tip SHV prin metoda difuzimetrica
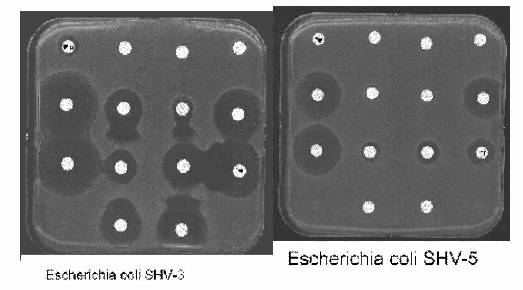
Fig. 2 Aspectul antibiogramei la tulpini de E. coli producatoare de ß-lactamaze de spectru larg:
stg. aspecte de sinergism (original).
Principiu: Aceasta metoda se bazazeaza pe proprietatea ESBL de a fi sensibile la inhibitori de β-lactamaze (acidul clavulanic). Exista si exceptii de la aceasta regula enzimele de tip IRT (Inhibitor Rezistant TEM βlactamase).
Benzile E-test contin un gradient de Cefotaxim la un capat si un gradient de Cefotaxim si acid clavulanic la celalat capat. Concentratia de Cefotaxim este mai mica in cazul in care acesta este asociat cu acidul clavulanic conform tabelului 23:
Tabelul 23: Concentratiile de antibiotic pe strip-urile E-test
|
Cefotaxim (mg/L) |
Cefotaxim (mg/L) + ac. Clavulanic |
Benzile similare continand gradiente de ceftazidim / ceftazidim + clavulanat (TZ / TZL) sunt deasemenea disponibile si sunt mai potrivite pentru detectarea tipului enzimatic CTX-M.
Daca raportul dintre concentratia minima inhibitorie (CMI) de Cefotaxim si CMI de Cefotaxim + clavulanat este mai mare de 8 rezulta ca tulpina respectiva este producatoare de ESBL-uri.
Alternativ, se pot inregistra urmatoarele aspecte ale placii in urma incubarii:
Crestere bacteriana de-a lungul intregii benzi (lipsa elipsei de inhibitie) indica faptul ca valoarea respectiva de CMI este mai mare decat cea mai mare valoare din scala (Fig. 29

O elipsa de inhibitie dedesubtul gradientului arata o valoare a CMI mai mica decat
cea mai mica valoare din scala.
Aparitia unei zone fantoma plasata sub gradientul CT, desi o zona de inhibitie clara in jurul capatului CT nu s-a evidentiat, indica producerea de ESBL (Fig. 30).

Aparitia unei elipse de inhibitie deformate inspre capatul tiparit indica producerea de ESBL, datorita sinergismului dintre cefotaxim (CT) si acidul clavulanic difuzat dinspre regiunea CTL (Fig 31).
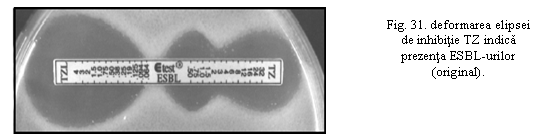

Fig. 32. Aspectul E-Test la o tulpina de E.coli J53 (SHV-2) producatoare de ESBL
(dupa L. Stratchounski, I si colab., 2003)
CMIAMC
12 mg/L
CMIPTZ 1,5 mg/L
CMICPS1 mg/L

Fig. 33. Aspectul E-Test la o tulpina de K.pneumoniae producatoare de ESBL (dupa L. Stratchounski, I si colab., 2003)
(SHV-2 + SHV-1 + TEM-1)
CMIAMC 48 mg/L
CMIPTZ 64 mg/L
CMICPS 4 mg/L
A.2. Metoda discurilor duble
Aceasta metoda presupune compararea diametrelor zonelor de inhibitie date de un disc impregnate cu o cefalosporina de spectru larg singura cu un alt disc impregnat cu acelasi antibiotic asociat cu un inhibitor de βlactamaze (acidul clavulanic). Daca tulpina testata produce o enzima de tip ESBL, diametrul zonei este mai mare pentru discul ce contine antibiotic + inhibitor. NCCLS recomanda compararea diametrelor zonelor de inhibitie date de catre cefotaxim 30 µg/disc cu cele ale cefotaximului + acid clavulanic 30 +10 µg/disc, precum si ceftazidim 30 µg/disc cu cele ale ceftazidimului + acid clavulanic 30 + 10 µg/disc. Mzali si colab., 2000 au evaluat discuri cu antibiotice si inhibitori tip MAST DD, folosind metode conform indicatiilor NCCLS, si au obtinut o sensibilitate de 93% a acestei metode pentru tulpini bacteriene control, producatoare de ESBLuri, in timp ce perechile de ceftazidim si cefotaxim s-au dovedit a avea o sensibilitate mai scazuta: 86% si respectiv 66%.
O varianta a metodei discurilor duble cu antibiotice (produse de OXOID) se bazeaza pe compararea diametrelor zonelor de inhibitie produse de cefpodoxim 10 µg/disc si cefpodoxim + clavulanat 10 + 1 µg/disc. Producerea de ESBL-uri este confirmata daca diametrul zonei de inhibitie data de discul ce contine si clavulanat este cu 5 mm mai mare decat diametrul zonei discului fara inhibitor (Carter si colab., 2000).
Testul dublei difuzii pentru detectarea ESBL
Pe o placa cu mediu Mueller Hinton inoculata cu tulpina de testat (cultura de 24 h, insamantare pe placa prin tehnica tamponului) se aplica discuri cu amoxicilina / clavulanat (AMC) si ceftazidim (CAZ) la 25 -30 mm distanta (Jarlier si colab., 1988 ). De cealalta parte a discului AMC se plaseaza un disc continand o alta cefalosporina, de preferat cefotaxim (CT ).
Se incubeaza peste noapte la 37oC.
Producerea de ESBL este certificata daca diametrul zonei de inhibitie este extins din cauza clavulanatului ( inhibitorul de β-lactamaze) (Fig 34, 35).
Avantajul acestei metode simple este costul scazut; dezavantajul este acela ca separarea optima a discurilor poate varia cu fiecare tulpina. Izolatele cu ESBL-uri de tip TEM si SHV dau rezultate pozitive, in schimb cele cu enzime de tip CTX-M dau un rezultat pozitiv numai atunci cand cefotaximul sau cefpodoximul inlocuiesc ceftazidimul, ca indicator de cefalosporine.
Tulpinile producatoare de ApmC si majoritatea tulpinilor hiperproducatoare de enzime K1, dau rezultate negative, cu toate cele trei cefalosporine.
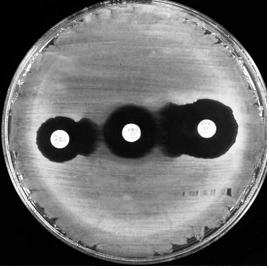
Fig. 34. Detectarea producerii de ESBL prin testul dublei difuzii.
Discuri cu antibiotice: stanga: CAZ, centru: AMC, dreapta: CT (original).
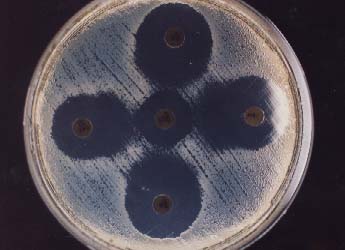
Fig. 35. Detectarea ESBL prin testul dublei difuzii AMC-amox. si acid clavulanic in centru; . CAZ-Ceftazidim. ATM-Aztreonam, CTX-Cefotaxim, CRO-Ceftriaxone (dupa Shah si colab., 2003).
Se recomanda utilizarea discurilor de cloxacilina pentru detectarea penicilinazelor (acest antibiotic inhiba cefalosporinazele);
N.B. inoculul dens poate masca hiperproductia de beta-lactamaze;
N.B.- cand sinergismul nu este vizibil din cauza fuzionarii zonelor de inhibitie, se indeparteaza discurile (30 mm) sau se taie discurile de antibiotic in patru.
N.B.- existenta unor discrepante mari intre diametrul zonelor de inhibitie la CAZ si CTX se datoreaza prezentei unei alte enzime pe langa ESBL (ex. β-lactamaze AmpC la Enterobacter cloacae).
Teste pentru confirmarea prezentei -lactamazelor cromosomale inductibile ( -lactamaze tip AmpC)
Cefalosporinele de generatie I (ampicilina si amoxicilina) induc exprimarea enzimelor tip AmpC si sunt distruse de catre acestea, la majoritatea speciilor de Enterobacteriaceae. Consecutiv, tulpinile bacteriene producatoare de β-lactamaze inductibile tip AmpC devin rezistente constitutiv. Speciile producatoare de β-lactamaze inductibile tip AmpC segrega mutante derepresate care produc enzime AmpC fara inductie (derepresie stabila). Aceste mutante sunt rezistente la aproape toate penicilinele si cefalosporinele (Livermore si colab, 2001) si sunt de asemeni frecvente in izolatele clnice.
Speciile producatoare de β-lactamaze tip AmpC pot fi recunoscute prin teste de antagonism intre cefoxitim si cefotaxim (Sanders si colab. 1982). Pentru efectuarea acestui test se inoculeaza placi, conform protocolului din metoda antibiogramei, si se plaseaza discuri cu ceftazidim si cefoxitim, pe acceasi placa, la o distanta de 25 mm. Inductia de β-lactamaze este evidentiata prin aplatizarea zonei de inhibitie a ceftazidimului in dreptul discului de cefoxitim. Astfel, cefoxitimul a determinat producerea de β-lactamaze iductibile care au hidrolizat ceftazidimul (Fig 36).
Acidul clavulanic este un inductor al β-lactamazelor tip AmpC si de aceea aparitia unor mici colonii in interiorul zonei de inhibitie a discului AMC (amoxicilina - acid clavulanic) poate fi un indicator util pentru detectarea prezentei enzimelor AmpC.
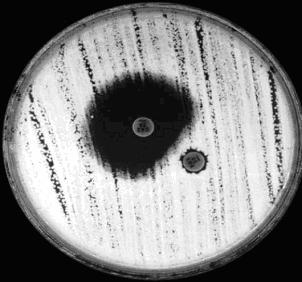
Fig. 36: Detectarea β-lactamazelor inductibile tip AmpC. Discul cu diamterul zonei de inhibitie mare este ceftazidim (CAZ). Cel cu diamtrul foarte mic este cefoxitim (FOX) (original).
Au fost concepute numeroase teste directe de detectare a activitatii β-lactamazice, dar numai o parte se preteaza ca analize de rutina. Majoritatea folosesc cefalosporine cromogene, sau coreleaza hidroliza penicilinei cu o schimbare a culorii mediului de reactie, mediata de iod sau detectata cu un indicator de pH.
Cefalosporinele cromogene sunt foarte specifice, in timp ce acidifierea mediului si reducerea iodului se poate datora si altor cauze decat actiunii β-lactamazelor, generand rezultate fals pozitive. Datorita slabei specificitati a testului iodometric ar trebui realizate in paralel controlale pozitive si negative pentru toate testele.
Testul la nitrocefin
Nitrocefinul este o cefalosporina care poate functiona ca substrat cromogen pentru β-lactmaze, si isi schimba culoarea de la galben la rosu atunci cand este hidrolizat. Acesta reprezinta testul cel mai sensibil pentru majoritatea βlactamazelor, exceptand penicilinazele stafilococice. Avantajul folosirii acestui antibiotic in testele de evidentiere consta in faptul ca el este hidrolizat de toate β lactamazele cunoscute, indiferent de specificitatea acestora.
Nitrocefinul este disponibil sub forma de pudra purificata de la Becton Dickinson (Oxford, UK) sau discuri imbibate cu antibiotic (OXOID).
Coloniile bacteriene sunt raclate de pe mediul nutritiv solid si resuspendate in tampon fosfat obtinandu-se o suspensie densa, peste care se adauga solutie nitrocefin. Activitatea β-lactamazica este evidentiata prin aparitia culorii rosii in 1 - 2 minute. In cazul enzimelor cu activitate mai mica raspunsul poate intarzia, insa rezultatele pozitive aparute la mai mult de 10 minute trebuie tratate cu scepticism intrucat ele pot fi cauzate de o activitate βlactamazica secundara a proteinelor care leaga penicilina (penicillin binding protein - PBP) care formeaza complexe acil instabile (OCallaghan si colab., 1972).
Avantajul acestei metode consta in rapiditatea cu care se pot obtine rezultate despre prezenta β-lactamazelor.
Testul iodometric
Hidroliza penicilinei determina formarea acidului peniciloic, care reduce iodul, decolorand complexul iodamidon. Aceasta proprietate poate fi exploatata pentru a detecta activitatea βlactamazica in tuburi sau pe hartie. Aceste teste sunt sensibile pentru penicilinazele stafilococice, dar sunt mai putin sensibile decat nitrocefinul pentru majoritatea βlactamazelor provenite de la bacterii Gram negative.
Metoda in tub: Se poate efectua in cantitati mici atat in tuburi, cat si in placi cu godeuri. Se distribuie benzilpenicilina (penicilina G) in tampon fosfat, in godeurile unei placi. In aceasta solutie se suspenda cultura bacteriana, solida (de pe agar) si se pastraza la temperatura camerei pentru aproximativ 30 60 minute. Se adauga amidon solubil in apa distilata si ulterior iod in solutie de iodura de potasiu. Activitatea β lactamazica este demonstrata prin decolorarea iodului in maxim 5 minute (Catlin, 1975). Sunt necesare teste de control pozitive si negative deoarece alte proteine pot de asemenea reduce iodul, iar un inocul concentrat poate da rezultate fals pozitive.
Metoda pe benzi de hartie: Se foloseste hartie de filtru Whatman nr.3, imbibata in solutie de amidon dizolvat in apa distilata la fierbere, la care s-a adaugat benzil penicilina la racire. Cand benzile de hartie iodometrica s-au uscat (aproximativ 2 ore), acestea sunt inmuiate in solutie de iod in iodura de potasiu. Pe acestea se aplica colonii dintr-o cultura de 24 de ore. Decolorarea apare in 5 minute si indica activitate β lactamazica (Jorgensen si colab. , 1977).
Testul acidimetric
Hidroliza nucleului β-lactamic genereaza o grupare carboxil care acidifica un mediu netamponat. Acidifierea rezultata poate fi detectata in tuburi sau pe hartie de filtru.
Metoda in tub: Se foloseste solutie de rosu fenol la care s-a adaugat benzilpenicilina, cu pH 8,5. Solutia de culoare violet este distribuita in godeuri si inoculata cu bacterii din cultura solida pentru a forma o suspensie densa. Activitatea β-lactamazica este indicata de virajul culorii de la rosu la galben in maxim 5 minute (Duma & Kinz, 1968).
Metoda pe benzi de hartie: benzi mici, de hartie de filtru Whatman sunt imbibate intr-o solutie proaspat preparata de benzilpenicilina, bromo-crezol si NaOH. Benzile se usuca si se pastreaza pentru mai multe luni. Inainte de folosire ele trebuiesc rehidratate in apa distilata si apoi se aplica cultura bacteriana de pe agar solid. Aparitia culorii galbene in maxim 5 minute indica activitate β-lactamazica (Livermore, 1995).
A.3. Metode de identificare a diferitelor tipuri de β-lactamaze: cinetica enzimatica
Obtinerea extractului proteic
Tulpinile bacteriene care expima constitutiv β-lactamaze se cultiva in bulion Mueller-Hinton peste noapte la 37oC, din care se recolteaza celulele prin centrifugare. Suspensia celulara obtinuta din fiecare preparat se supune sonicarii urmata de ultracentrifugare. Supernatantul este dializat si concentrat daca este nevoie (Iaconis si colab., 1989).
Purificarea β-lactamazelor
Intr-o prima etapa, β-lactamazele se purifica prin focusare isoelectrica orizontala (IEFO), intr-un gradient de pH cuprins intre 6.5 si 10.5 obtinut prin metoda amfolitilor.
Dupa migrare, gelul este impartit in 30 de segmente egale in care se testeaza activitatea βlactamazica cu nitrocefin sau benzilpenicilina (OCallaghan si colab. , 1972). Fractiile din gel care prezinta activitate β lactamaizica se incarca pe o coloana cromatografica de carboxi metil Sephadex si se elueaza cu tampon fosfat conform protocolului descris de Iaconis si colab., 1989. Fractiile care au prezentat activitate β lactamazica se concentreaza prin ultrafiltrare si au fost trecute in continuare pe o coloana schimbatoare de ioni.
Hidroliza antibioticelor β-lactamice este examinata prin spectrofotometrie UV (Beckman DU-7). Se folosesc cuve de cuartz, de 1cm, la lungimea de unda maxima de absorbtie a inelului β-lactam, pentru fiecare antibiotic in parte, iar densitatea optica se masoara la intervale de 10 secunde, timp de 5 minute.
Maximul de absorbtie pentru pricipalele antibiotice folosite sunt:
cefalotin (265 nm)
cefuroxim (274 nm)
cefotaxim (254 nm)
ceftazidim (254 nm)
imipenem (299 nm)
cloxacilin (260 nm)
aztreonam (292 nm)
nitrocefin (489 nm)
Rata de hidroliza a benzilpenicilinei este masurata la 233 nm. Solutiile de antibiotic sunt preparate in tampon 0,1M, pH 7.0. Pentru stabilirea profilelor substratelor, toate antibioticele se examineaza la o concentratie de 100 uM, mai putin benzilpenicilina care este testata pentru 500 uM.
O unitate de activitate β-lactamazica, este definita ca fiind cantitatea de enzima care hidrolizeaza un nmol de substrat / min in etapa liniara a reactiei la 37oC in tampon fosfat 0,1M, ph 7.0.
Constanta Michaelis (Km) si rata maxima de hidroliza (vm) se determina cu ajutorul transformarii Lineweaver-Burk a vitezei initiale (v) pentru sase concentratii diferite ale substratului. Susceptibilitatea β-lactamazelor de a fi inhibate de clavulanatul de potasiu si cloxacilina este masurata cantitativ prin preincubarea enzimei cu concentratii diferite de inhibitor pentru 10 minute. Pentru masurarea activitatii enzimatice reziduale se adauga ulterior ca substrat benzilpenicilina (500 uM) sau imipenem (100 uM).
Concentratia de inhibitor necesara pentru a inhiba 50% din activitatea enzimatica se determina prin teste statisitice de certitudine (Iaconis si colab., 1989).
Evidentierea fenotipica a rezistentei la aminozide la cocii Gram-pozitivi
La S. aureus rezistenta la aceste antibiotice este enzimatica fiind evidentiate mai multe fenotipuri (tabelul 24).
Tabelul 24: Rezistenta S. aureus la aminozide
|
Fenotip |
Enzime |
Kanamicina Amikacina Isepamicina |
Tobramicina |
Gentamicina Netilmicina |
|
salbatic |
S |
S |
S |
|
|
K |
APH3-III |
R |
S |
S |
|
KT |
ANT-4-4 |
R |
R |
S |
|
KTG |
APH 2-AAC6 |
R |
R |
R |
APH = aminozid-fosfotransferaza; ANT = aminozid nucleotidiltransferaza; AAC = aminozid acetiltransferaza
Rezistenta la aminoglicozide la enterococi
Rezistenta naturala de nivel scazut (intrinseca) se datoreaza unui transport ineficient al acestora prin membrana citoplasmatica, insa activitatea lor este sinergica cu β-lactaminele si glicopeptidele. Specia E. faecium produce in mod natural o acetiltransferaza cromosomala AAC(6) care inactiveaza aminozidele gentamicina ramane activa).
Un numar din ce in ce mai mare de tulpini de enterococi manifesta rezistenta de nivel inalt la aminoglicozide (HLR) - definita ca CMI ≥2 mg/mg si, concomitent, rezistenta la sinergismul cu beta-lactamicele (Tabelul 25). Mecanismul de rezistenta preponderent este modificarea enzimatica a aminoglicozidelor, ca urmare a achizitionarii uneia sau mai multor gene. Enzimele codificate de aceste gene sunt sintetizate constitutiv si au localizare citoplasmatica. Modificarea enzimatica a antibioticului este asociata cu o diminuare a afinitattii sale pentru proteinele ribosomale (Shaw si al., 1993) si cu alterarea transportului antibioticului.
Tabelul 25: Principalele fenotipuri de rezistenta ale enterococilor la aminozide
|
Enzime |
Kanamicina |
Gentamicina |
Netilmicina |
Tobramicina |
Amikacina |
|
S |
S |
S |
S |
S |
|
|
APH3 |
R |
S |
S |
S |
R |
|
ANT-4 |
R |
R |
S |
R |
S/R |
|
APH2-AAC6 |
R |
R |
R |
R |
R |
APH = aminozid fosfotransferaza; ANT = aminozid nucleotidiltransferaza; AAC = aminozid acetiltransferaza
Enterobacteriile cu exceptia speciei Providencia stuartii, sunt in mod natural sensibile la aminoglicozide. Cele patru aminozide utilizate in terapia infectiilor cu enterobacterii sunt: gentamicina (GM), tobramicina (TM), netilmicina (NET) si amikacina (AN).
P. stuartii are o rezistenta cromosomala la gentamicina, netilmicina si tobramicina. Rezistenta dobandita la aceste antibiotice este putin frecventa (tabelul 26).
Anumite fenotipuri de rezistenta sunt generate in functie de prezenta anumitor enzime care modifica aminozidele:
Fenotipul 1: tulpini rezistente la gentamicina (GM), asociate cu prezenta unei 3-aminoacil-acetiltrensferazei AAC;
Fenotipul 2: tulpini rezistente la gentamicina (GM) si la netilmicina (NET) asociate cu o 2-nucleotidiltransferaza ANT;
Fenotipul 3: tulpini rezistente la tobramicina (TM), netilmicina (NET), si la amikacina (AN) asociate cu o 6-aminoacilacetiltransferaza AAC.
Tabelul 26: Fenotipurile de rezistenta dobandita ale enterobacteriilor la aminozide
|
Fenotip |
Enzime |
Gm(G) |
To(T) |
Nt |
Ak(A) |
Is(I) |
Specii implicate |
|
G |
AAC(3)-I |
R |
S |
S |
S |
S |
E.coli, P.mirabilis, K.pneumoniae, E. cloacae |
|
A |
APH(3)-VI |
S |
S |
S |
R |
R |
E. cloacae |
|
GT |
AAC(3)-VI |
R |
R |
S |
S |
S |
E.coli |
|
TA |
ANT(4)-II |
S |
R |
S |
R |
R |
TOATE |
|
GTNt |
AAC(2)-I AAC(3)-IV |
R R |
R R |
R R |
S S |
S S |
P.stuartii E.coli |
|
KTG |
ANT(2)-I |
R |
R |
S |
S |
TOATE |
|
|
KTANt |
AAC(6)-I |
S |
R |
R |
R |
S |
S.marcescens E.coli, E.cloacae K. pneumoniae E. aerogenes |
|
KTGNt |
AAC(3)-II |
R |
R |
R |
S |
S |
TOATE |
|
KTGANt |
Impermeabilitate Asociatie de enzime |
R R |
R R |
R R |
R R |
R R |
TOATE TOATE |
Gm = gentamicina, To = tobramicina, Nt = netilmicina, Ak =amikacina, Is =isepamicina, AAC = aminozid acetiltransferaze, ANT = aminozid nucleotidiltransferaza, APH = aminozid fosfotransferaze In fiecare clasa, enzimele sunt desemnate prin tipul lor de activitate, numarul carbonului purtand gruparea modificata (cifre arabe de la 1 la 6) si ciclul afectat de modificare (primar, secundar sau deloc). Cifra romana care urmeaza indica spectrul de substrat al enzimei (adica ansamblul de molecule hidrolizate).
Evidentierea fenotipurilor de rezistenta la aminoglicozide la Pseudomonadaceae
Fenotipul salbatic este sensibil la toate aminozidele.
Fenotipurile de rezistenta dobandita se datoreaza mai multor mecanisme: impermeabilitate, eflux, inactivare enzimatica, mutante respiratorii, combinatii ale acestor mecanisme (tabel ul 27, 28). La P. aeruginosa cel mai intalnit mecanism este cel de impermeabilitate (tabelul 27).
Tabelul 27: Principalele fenotipuri de rezistenta ale P. aeruginosa la la aminozide
|
Fenotip |
Enzima |
G |
T |
Nt |
A |
I |
|
G |
AAC(3)-I |
R |
S |
S |
S |
S |
|
GNt |
AAC(3) |
R |
S |
R |
S |
S |
|
G T |
ANT(2)-I |
R |
R |
S |
S |
S |
|
GNtT |
AAC(6)-II AAC(3)-II,IV,V |
R |
R |
R |
S |
S |
|
TNtA |
AAC(6 )-I |
S |
R |
R |
R | |
|
GTNtA |
AAC(6)+ANT(2)-I |
R |
R |
R |
R | |
|
Eflux(30-40%) |
R |
S |
R |
R |
R |
G = gentamicina ; T= tobramicina ; Nt =netilmicina ; A =amikacina ; I = isepamicina
AAC = amioacid-acetiltransferaza ; ANT = aminoacid- nucleotidiltransferaza.
Tabelul 28: Fenotipuri de rezistenta la aminozide la Acinetobacter spp
|
Fenotip de rezistenta |
Enzime |
Prevalenta ( %) |
|
G |
AAC(3)-I | |
|
GN |
AAC(3)-? | |
|
GTN |
AAC(3)- II AAC(3)- V | |
|
TNA |
AAC(6)-I | |
|
GTN |
AAC(6)- II | |
|
GT |
ANT(2 )-I | |
|
APH(3)-I | ||
|
AI |
APH(3)-VI |
G = gentamicina; N = netilmicina; T = tobramicina;
A = amikacina; I = isepamicina; APH = aminozidfosfotransferaza; ANT =
aminozidnucleotidiltransferaza; AAC = aminozidacetiltransferaza.
Evidentierea fenotipurilor de rezistenta la quinolone
Stafilococii sunt natural rezistenti la quinolone de prima generatie, in schimb sunt sensibili la fluorochinolone.
Sunt descrise 2 mecanisme de rezistenta:
afinitate scazuta prin modificarea tintei, ADN giraza. Dobandirea rezistentei este legata de aparitia mutatiilor cromosomale in genele gyrA si parC care provoaca o diminuare a legarii quinolonelor la tintele lor intracelulare, complexele ADN giraza si ADN topoizomeraza 4.
eflux
Pneumococii prezinta rezistenta naturala la quinolonele din prima generatie si au sensibilitate redusa la pefloxacin, ofloxacin si ciprofloxacin. Foarte active pe pneumococ sunt levofloxacina si moxifloxacina.
Mecanismele rezistentei dobandite sunt: efluxul activ si mutatiile topoizomerazei IV (gena parC) si girazei (gena gyrA)
Toate enterobacteriile prezinta rezistenta incrucisata la toate chinolonele, dar nivelul de exprimare variaza pentru fiecare substanta antimicrobiana in functie de bacterie. Pentru a pune in evidenta diferite fenotipuri se testeaza acidul nalidixic (NAL) si doua florochinolone: de exemplu: pefloxacina (PEF) sau ofloxacina (OFX) si ciprofloxacina (CIP) sau norfloxacina (NOR) (tabelul 29).
Au fost identificate urmatoarele fenotipuri de rezistenta la chinolone la enterobacterii:
Fenotipul 1: tulpini sensibile la toate chinolonele;
Fenotipul 2: tulpini rezistente la acid nalidixic, dar sensibile la fluorchinolone;
Fenotipul 3: tulpini cu sensibilitate diminuata la pefloxacina si la ofloxacina, dar sensibile la ciprofloxacina.
Fenotipul 4: tulpini rezistente la toate chinolonele.
Tabelul 29: Fenotipuri de rezistenta a enterobacteriilor la chinolone
|
Fenotipuri |
Ac. nalidix (1) |
Norfloxacin |
Pefloxacin |
Ofloxacin |
Ciprofloxacin |
|
I |
S |
S |
S |
S |
S |
|
II |
R |
S |
S |
S |
S |
|
III |
R |
I/R |
I/R |
S |
S |
|
IV |
R |
R |
R |
R |
R |
|
Rar eflux (E. coli) |
S |
R |
S |
S |
S |
(1) Ac. Nalidix si toate chinolonele clasice (prima genetatie).
Fenotipurile de rezistenta la chinolone la P. aeruginosa
Fenotipul salbatic este sensibil la toate fluorochinolonele desi in terapie cel mai utilizat este ciprofloxacinul (tabelul 30). Rezistenta dobandita se datoreaza urmatoarelor mecanisme: impermeabilitate (porine si LPS); modificarea afinitatii tintei (subunitatile A si B ale ADN-girazei si Cssi D ale topoizomerazei IV; Eflux activ (OprM, OprJ, OprN confera rezistenta de nivel scazut)
Tabelul 30: Principalele fenotipuri de rezistenta ale P. aeruginosa la chinolone
|
Fenotip |
Nofloxacin |
Pefloxacin |
Ofloxacin Levofloxacin |
Ciprofloxacin |
|
I (salbatic) |
S |
S |
S |
S |
|
II |
I |
I |
I |
S |
|
III |
R |
R |
R |
S |
|
IV |
R |
R |
R |
R |
|
eflux |
R |
S |
S |
R |
Se considera ca aproape 70% din tulpinile de A. baumanii au dobandit rezistenta incrucisata la fluoroquinolone, prin mutatii la nivelul tintei.
Cazuri de rezistenta sporadica la glicopeptide au fost descrise la stafilococii coagulazo-negativi inca din 1981. In primele cazuri rezistenta la vancomicina si teicoplanina a fost de nivel scazut (CMI 8-16mg/ml). Tulpinile raportate ulterior aveau CMI intre 16 si 64 mg/ml la teicoplanina si o sensibilitate mentinuta la vancomicina. Fenotipul de rezistenta a fost asociat cu prezenta unei proteine membranare de 35 000-39 000da la S.epidermidis si S.haemolyticus.
Cateva tulpini sunt intermediare la vancomicina (tulpini GISA) si au o rezistenta crescuta la teicoplanina. Acestea au fost izolate de la pacienti care au urmat multiple tratamente cu vancomicina. Aceste tulpini GISA nu au genotipul VanA, VanB, VanC, mecanismul de rezistenta fiind inca necunoscut. Articole recente raporteaza tulpini de S.aureus cu sensibilitate redusa la teicoplanina (cresterea CMI de 8 ori) si la vancomicina (CMI dublu), tulpini care sintetizeaza o noua proteina de 39 000da (Fig. 36).
In schimb, tulpini de S.aureus sensibile la vancomicina si cu sensibilitate redusa la teicoplanina poseda o proteina membranara de 35 000da si prezinta o crestere a exprimarii PLP 2-1 si PLP 2-2.
Rezistenta la glicopeptide este uneori greu de depistat prin antibiograma difuzimetrica. Comitetul de Antibiograma al Societatii Franceze de Microbiologie (CA-SFM) recomanda determinarea CMI pentru toate tulpinile al caror diametru de inhibitie este mai mic de 17 mm. Detectarea rezistentei pare sa dea rezultate mai bune pe mediu semi-gelozat.
Rezistenta enterococilor la vancomicina
Rezistenta enterococilor la vancomicina a inceput sa fie semnalata din anul 1988 (Leclercq si al., 1988 & 1989). Supravegherea clinica, biochimica si genetica a rezistentei la glicopeptide in cadrul genului Euterococcus a evidentiat o mare heterogenitate fenotipica si genotipica. Cele trei fenotipuri (VanA. VanB & VanC) sunt definite prin nivelul de rezistenta la vancomicina si sensibilitate sau rezistenta la teicoplanina.
Rezistenta intrinseca (naturala) la glicopeptide numita VanC se caracterizeaza prin rezistenta de nivel scazut la vancomicina si sensibilitate la teicoplanina intalnita la speciile: E. gallinarum, E. casseliflavus si E. flavescens. Gena VanC codifica o ligaza care substituie D-Ala-D-Ser in locul D-Ala-D-Ala ca precursor al peptidoglicanului.
Mecanismul rezistentei dobandite este legat de producerea unui nou precursor al peptidoglicanului in care dipeptidul terminal ( D-Ala-D-Ala) este inlocuit de D-Ala- D-Lac, a carui afinitate pentru glicopeptide este redusa. Sunt descrise sase fenotipuri de rezistenta dobandita: VanA, VanB, VanD, VanE,VanG, Van E (la E. faecalis si E. faecium).
|
Fenotip |
VanA (HLR) |
VanB (MLR) |
VanC |
VanD |
VanG |
VanE |
|
CMI Va(mg/l) |
64->1024 | |||||
|
CMI Tec(mg/l) | ||||||
|
Expresie |
Inductibila |
Inductibila |
Constitutiva |
Constitutiva |
Inductibila |
Inductibila |
|
Suport genetic |
Plasmidial transpozon Tn1546 |
Cromosomal |
Cromosomal |
Cromosomal |
Cromosomal |
Cromosomal |
|
Transferabil prin conjugare | ||||||
|
Specii bacteriene |
E. faecium E. faecalis E. gallinarum E. flavescens E.avium E. durans E. hirae |
E.faecalis E. faecium |
vanC1 - E. gallinarum, vanC-2 - E. casseliflavus, and vanC-3 - E. flavescens |
E.faecium |
E.faecium E. faecalis |
E. faecalis |
Transpozonul Tn1546 codifica 9 proteine:
doua necesare transpozitiei (ORF1 si ORF2)
doua implicate in reglarea expresiei inductibile a rezistentei ( VanR siVanS)
cinci implicate in rezistenta (VanH-dehidrogenaza, VanA-ligaza, VanX-peptidaza, majore si VanY si VanZ, considerate minore).
A fost descrisa o tulpina de S. bovis rezistenta la vancomicina care poseda gena vanB.
Cea de-a doua forma a rezistentei la vancomicina a rezultat probabil din transferul conjugativ al operonului vanA de la Enterococcus faecalis vancomicin-rezistent (fig. 37). Showsh si colaboratorii au demonstrat ca plasmidul de la Enterococcus faecalis care contine vanA codifica de asemenea un feromon del sex care este sintetizat si de catre S. aureus, sugerand posibilitatea transferului conjugal. Aceste izolate VRSA au rezistenta de nile inalt la vancomicina, cu CMI de 128 µg/ml. Rezistenta la aceste izolate este cauzata de catre alterarea peptidului terminal la D- Ala-D-Lac in loc de D-Ala-D-Ala (fig. 38). Sinteza D-Ala-D-Lac se produce doar prin expunerea la concentratii mici de vancomicina.
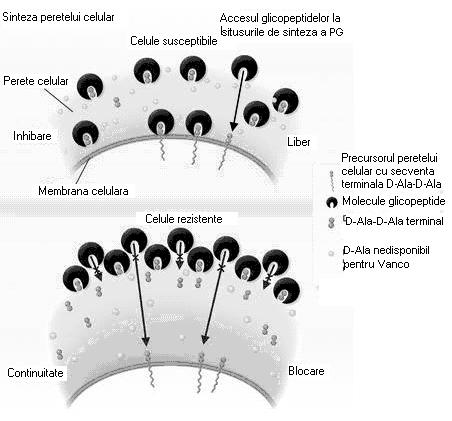
Fig. 37. Mecanismele de rezistenta la vancomicina a tulpinilor VISA. Acestea sintetizeaza in exces peptidoglican, avand ca rezultat cresterea numarului de resturi D-Ala-D-Ala care leaga vancomicina (dupa Walsh si Howe, 2002).
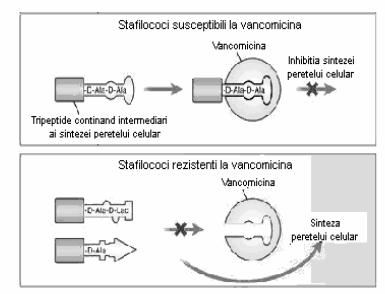
Fig. 38: Mecanismul de rezistenta la vancomicina a tulpinilor VRSA. (dupa Hiramatsu si colab., 1997).
Evidentierea rezistentei la macrolide, streptogramine, lincosamide (MLSb)
Trei mecanisme sunt implicate in rezistenta la aceste molecule:
1. Modificarea tintei ribosomale (sub-unitatea 50S)
Tulpinile rezistente produc o metilaza responsabila de o dimetilare specifica a adeninei din ARNr 23S. Aceasta provoaca o schimbare de conformatie a ARN, care reduce afinitatea MLS pentru ribosomi. Metilarea se produce la toate macrolidele si lincosamidele. Aceasta rezistenta depinde de diferiti determinanti genetici. La stafilococ gasim genele ermA, ermB, ermC. Aceste gene sunt purtate de transpozoni (ermA) sau plasmide (erm). Exprimarea fenotipica a rezistentei poate fi inductibila (rezistenta crescuta pentru toate macrolidele cu 14 si 15 atomi de carbon si sensibilitate la macrolidele cu 16 atomi de carbon), sau constitutiva (rezistenta crescuta la toate macrolidele cu 14-16 atomi de carbon, lincozamidele si streptograminele B rezistenta de tip MLSB) (tabelul 31). Rezistenta compusului B nu modifica sinergia a doi compusi ai streptograminelor.
|
Mecanism |
Genotip |
Fenotip |
14C1 15C2 |
16C3 |
LIN |
CLIN |
PRIB |
PRIA |
PRIi5 |
|
Modificarea tintei |
erm inductibil |
MLSB |
R |
S |
S |
S |
S |
S |
S |
|
erm constitutiv |
MLSB |
R |
R |
R |
R |
S |
S |
S |
|
|
Inactivare |
linA |
L |
S |
S |
R |
S4 |
S |
S |
S |
|
Vat/vatB |
(L)SA |
S |
S |
S |
S |
S |
R | ||
|
vgb |
SB |
S |
S |
S |
S |
R |
S | ||
|
Eflux |
vga |
(L)SA |
S |
S |
S/I/R |
R/I/S |
S |
R |
S/R |
|
msrA |
Erm |
R |
S |
S |
S |
S |
S |
S |
(1) 14C: eritromicina, roxitromicina, claritromicina, diritromicina
(2) 15C: azitromicina
(3) 16C: spiramicina, josamicina, midecamicina, tilosina
(4): antibiotic a carui activitate bacteriostatica sau bactericida este diminuata
(5) Pri : pristinamicina
2. Inactivarea antibioticului
Streptogramin A acetiltransferaza confera rezistenta la factorul A astfel incat streptogramin B hidrolaza, codificata de catre gena vgb, inactiveaza factorul B. Cel putin 2 gene vat si vatB sunt responsabile pentru acetilarea streptograminelor A. Rezistenta izolata la lincosamide, exceptionala la S.aureus dar detectata la stafilococii coagulazo-negativi (S.sciurii, S.cohnii, S.xylosus), este datorata achizttionarii unei enzime care modifica prin adenilare lincomicina si clindamicina. Fenotipul LSA poate fi confundat cu fenotipul L, ambele rare la S.aureus. Lincosamidele sunt inactivate de catre nucleotidtransferaze codificate de catre 2 gene plasmidiale foarte apropiate, linA si linA.
3. Efluxul activ de molecule
Exprimarea acestei rezistente depinde de gena msrA, care codifica pentru proteina MsrA responsabila de eliminarea crescuta a eritromicinei intrabateriene. Au fost izolati stafilococi rezistenti doar la streptogramina A. Rezistenta dependenta de gena vga este datorata unei diminuari intracelulare a antibioticului.
La enterococi, rezistenta naturala se manifesta la lincosamide si la compusul A al streptograminelor (la toate speciile cu exceptia E. faecium si E. durans, astfel ca acest tip de rezistenta poate fi utilizat ca test de orientare pentru identificare).
Rezistenta dobandita se manifesta prin doua mecanisme: modificarea tintei si inactivarea antibioticului.
Modificarea tintei (subunitatea 50S ribosomala) este mecanismul principal care explica rezistenta incrucisata la MLSK. La enterococi gena ermB este responsabila de aceasta rezistenta.
Inactivarea antibioticului se manifesta la E. faecalis prin actiunea unei lincomicinnucleotidiltransferaze (codificate de gene linA) care confera rezistenta la streptograminele A.
Rezistenta dobandita a pneumococului la macrolide este MLSB, datorita metilarii ARNr 23S, care diminueaza afinitatea macrolidelor pentru tinta. Aceasta rezistenta este incrucisata pentru toate macrolidele, lincosamidele si streptograminele B si poate fi de tip inductibil sau constitutiv.
Tulpinile care au gena erm imductibila sunt sensibile la ketolide la fel ca cele rezistente prin mecanismul de eflux .
Alte tulpini, care poseda gena mefE, codifica o rezistenta prin mecanism de eflux ele fiiind rezistente la eritromicina, dar sensibile la clindamiciaa, quinupristina si la ketolide. Genele erm si mefE sunt rareori asociate.
Al treilea fenotip de rezistenta a fost descris la un grup de 10 tulpini care nu au genele erm sau mefE dar sunt ery-R constitutiv si clinda-R.
La S.pyogenes mai putin de 10% dintre tulpini sunt rezistente la eritromicina, cu o mare heterogenitate
de la o zona la alta (18,3% in Asia, 10,6% in Europa de Est, 9,8% in Europa de Vest).
Doua mecanisme sunt responsabile de aceasta rezistenta: modificarea tintei, care da fenotipul MLSB constitutiv, codificat de gena ermB si un mecanism de efux care da fenotipul M codificat de gena mefA.
Telitromicina are o activitate intrinseca superioara celei a macrolidelor.
Evidentierea fenotipurilor de rezistenta la tetracicline
Sunt descrise 2 mecanisme de rezistenta la tetracicline:
primul este legat de o insuficienta concentrare intracelulara a antibioticului datorita unui eflux excesiv al antibioticului prin membrana citoplasmatica, sub actiunea unei proteine membranare TET. Acest mecanism confera o rezistenta la tetraciclina, dar nu si la minociclina.
al doilea mecanism este datorat unei modificari a tintei ribosomale a acestor antibiotice. In acest caz, exista o rezistenta crescuta la toate tetraciclinele.
La streptococi si enterococi au fost evidentiate 3 gene de rezistenta la tetraciclina, si anume: tetM, tetO (Zilhao si colab., 1988) si tetL (Chopra si colab., 1992).
Rezistenta la trimetoprim-sulfometoxazol
Rezistenta stafilococului la acesti 2 compusi se realizeaza prin mai multe mecanisme:
- modificarea enzimelor tinta (DHFR si THFR)
- hiperproducerea acestor enzime
Rezistenta la fosfomicina
Rezistenta stafilococului la fosfomicina este rezultatul unor mutatii foarte frecvente ce tintesc ransportul de antibiotic. A fost raportata o rezistenta plasmidiala la tulpini de stafilococ coagulazo-negativi si la S.aureus (gena fosB la S.aureus). Mecanismul, prin analogie cu cel descris la bacilii Gram negativi, ar putea fi inactivarea antibioticului prin conjugare cu glutation. Specia saprophyticus este natural rezistenta la fosfomicina.
Rezistenta la acid fuzidic
Rezistenta stafilococului la acidul fusidic este datorata unor mutatii care au loc cu frecventa mare, fapt pentru care in clinica acest antibiotic se utilizeaza numai in asociere cu alte antibiotice.
Rezistenta la rifampicina
Se disting doua niveluri de rezistenta:
- de nivel inalt cu CMI > 32mg/l
- sensibilitate diminuata (CMI 1- 4mg/l), tulpini izolate rar
Mecanismele de rezistenta la S.aureus nu sunt inca cunoscute.
La pneumococ, rezistenta la rifampicina, este legata de o mutatie a lantului β al ARN polimerazei care impiedica fixarea antibioticului pe tinta sa.
Rezistenta la cloramfenicol
Este predominant plasmidiala si se datoreaza prezentei la enterococi a unor gene cat omologe cu cele de origine stafilococica catpC221, catpC194 si catpSCS7 (Trieu-Cuot et al., 1993). CatpC221, este o gena, cu expresie inductibila. cu secventa cunoscuta (Shaw si al.1985), identificata pe plasmidul pC221 din S.aureus. Plasmidul conjugativ pIP501 (30,2 kb), cu spectru larg de gazda, a fost izolat pentru prima data din tulpina B96 de S. agalactiae (Horodniceanu si al., 1976). Prezenta sa confera tulpinii gazda fenotip Cmr-Emr. Studii bazate pe hibridizari ADN-ADN au evidentiat diseminarea determinantului catpIP501 pe plasmide izolate din enterococi (Pepper si al.,986).
Spre deosebire de stafilococi si enterococi unde este codificata plasmidial, la pneumococi cloramfenicol acetil transferaza este de natura cromosomala.
Rezistenta la oxazolidinone
La stafilococi, pana in prezent nu a fost evidentiata rezistenta incrucisata intre linezolid si alte antibiotice inhibitoare ale sintezei proteice, mai mult oxazolidinonele raman active asupra tulpinilor de MRSA.
Oxazolidinonele s-au dovedit a fi active asupra tulpinilor de E. faecalis si E. faecium fie ca sunt sensibile fie ca nu la glicopeptide. In vitro s-au obtinut mutante rezistente cu o frecventa de 10-9- 10-11 ( mutatii la nivelul subunitatii 23S ribosomale).
La penumococi nu s-a semnalat pana in prezent nici o tulpina rezitenta indiferent de sensibilitatea la penicilina G si macrolide.
6. Fenotipuri de rezistenta la alte microorganisme
B. cepacia este rezistenta, de asemenea, la aminozide, fosfomicina si polimixine, dar sensibila la fluoroquinolone si cotrimoxazol si, intr-un anumit procent, la carbapeneme in special la meropenem. Fenomenul de eflux inactiveaza asupra cloramfenicolul, trimetoprimul si ciprofloxacinul.
H. influenzae este rezistent la lincosamide si moderat sensibil la macrolide si ketolide. La aminozide sensibilitatea este variabila iar fata de cloramfenicol, tetracicline, rifampicina, quinolone si cotrimoxazol se manifesta o rezistenta de nivel scazut.
Biologul sau medicul de laborator care executa antibiograma are obligatia de a prezenta clinicianului rezultatul sub forma unui document usor de citit si de inteles, care trebuie sa urmeze logic rezultatului examenului microscopic direct si al culturii.
Mentiunile obligatorii din buletinul de analiza al antibiogramei sunt: metoda utilizata, lista antibioticelor conform DCI-urilor (denumire comuna internationala), raspunsul interpretat ca sensibil , intermediar , rezistent, CMI uri in anumite cazuri (in special pentru microorganismele izolate din infectii severe), mentionarea anumitor fenotipuri de rezistenta si consecintele lor terapeutice. In anumite cazuri este mai avantajos ca biologul sau medicul de laborator sa prezinte clinicianului doar rezultatele pentru anumite antibiotice, pentru a evita selectarea si emergenta unor fenotipuri de rezistenta in mediul spitalicesc (tabelul 33).
Antibiograma este inutila de fiecare data cand tulpina izolata nu poate fi considerata responsabila de infectia care a determinat prelevarea (ex. -microorganism comensal sau contaminant, cum este cazul streptococilor orali sau Haemophilus parainfluenzae in expectoratii, Bacillus intr-o singura hemocultura); cand numarul de UFC/ml este inferior pragului semnificativ (urina si expectorati); daca tulpina izolata poate fi considerata ca patogena, dar face parte dintre speciile sensibile in mod obisnuit la tratamentul de referinta si cand nu exista nici o corelatie in vitro-in vivo (ex. Legionella); in infectiile cutanate pentru care tratamentul este local sau chirurgical
Antibiograma trebuie practicata in acord cu clinicianul:
-Daca o bacterie comensala este izolata de la un pacient imunodeprimat;
- In prelevatele plurimicrobiene care contin specii aerobe/anaerobe (tabelul 32);
Tabelul 32: Criterii de decizie pentru realizarea unei antibiogramein functie de natura prelevatului
|
Prelevat |
Necesitatea antibiogramei |
Fara antibiograma |
Observatii |
|
Hemocultura |
Pentru toate speciile izolate cu exceptia contaminarii evidente |
Daca sunt contaminanti normali Bacillus, corineformi), iar o singura hemocultura este pozitiva | |
|
LCR |
Toate speciile considerate ca responsabile de meningite comunitare sau nosocomiale |
Daca sunt contaminanti normali Bacillus, corineformi SCN Daca examenul citochimic este normal |
S. pneumoniae / N. meningitidis: investigarea sistematica a rezistentei la penicilina G H.influenzae investigare β lactamaze Efectuare de CMI la antibioticele utilizate pentru tratament |
|
Cateter |
Daca numarul de colonii este ≥ 1000 ufc/ml (Brun Buisson) sau 15 colonii (Maki) |
Daca numarul de colonii este < 1000 UFC/ml ( Brun Buisson) sau 15 colonii ( Maki) | |
|
Urocultura |
Leucocituria ≥ 10000 UFC/ml si bacteriuria ≥ 100000 UFC/ml |
Daca leucocituria = 10000 UFC/ml si bacteriuria ≤ 1000 UFC/ml Daca > 3 specii bacteriene diferite |
Se realizeaza la: -pacientii imunodeprimat, dializati, diabetici, batrani, gravide, sau pacienti cu antecedente endoscopice sau chirurgicale pe caile urinare si leucociturie < 10000 si bacteriurie ≥ 100000 UFC/ml -la pacientii simptomatici si izolare de specii uropatogene sau pacientii cu antecedente de interventii endoscopice si chirurgicale pe caile urinare sau nefropatii subiacente si leucociturie ≥ 10000/ml si bacteriurie ≥ 1000 si < 100000 UFC/ml |
|
Secretii naso-faringiene |
Investigare de SAMR, Streptococ A la pacient alergic la penicilina | ||
|
Puroi din otite si sinuzite |
S. pneumoniae, Haemophilus influenzae, S. aureus, Enterobacterii, Pseudomonas |
Prelevate din otite externe, puroi recoltat prin paracenteza SCN |
S.pneumoniae investigarea rezistentei la penicilina |
|
Expectoratii |
≥10000000 UFC/ml nr. leucocite >25/camp nr. celule bucale < 10/camp (10x) |
Specii comensale sau patogene <10000000 UFC/ml |
Idem |
|
Secretii bronsice |
specii patogene ≥1000 UFC /ml |
<1000 UFC /ml |
H. influenzae (β lactamaza) |
|
Lichid bronho-alveolar |
specii patogene ≥10000 UFC /ml |
<10000 UFC /ml |
Idem |
|
Aspirate traheo-bronsice |
specii patogene ≥10000 UFC /ml |
<10000 UFC /ml |
Idem |
|
Coprocultura Coprocultura cantitativa la pacienti neutropenici |
Speciile responsabile de gastroenterite Toate speciile izolate ≥1000 UFC /ml |
Speciile comensale |
Interes epidemiologic in special |
|
Prelevate cutanate (panaritii, arsuri, ulcere varicoase,escare) |
biopsie cutanata arsi dermatomiozite |
escare profunde prelevare chirurgicala | |
|
Prelevate osteoarticulare |
Sistematic daca prelevarea e realizata chirurgical sau din situsuri profunde |
Fara antibiograma la orificii si fistule |
Conservarea tulpinilor timp de 1an |
|
Lichide de punctie (ascite, pleural) |
In functie de examenul microscopic si de nr. UFC |
Speciile comensale (SCN, corinebacterii) |
Se determina CMI Pentru antibioticele R |
|
Colectii inchise (abcese craniene si pulmonare) |
Cu prioritate pe speciile predominante | ||
|
Prelevate genitale |
N. gonorrhoeae sistematic -in functie de examenul microscopic nr. UFC pentru alte specii |
Gardnerella | |
|
Sperme |
UFC >1000 /ml | ||
|
Prelevate oculare |
Ulcere corneene, conjunctivite, endoftalmii |
Speciile comensale |
Tabelul 33: Agenti antimicrobieni de prima alegere pentru tratamentul infectiilor cu bacterii patogene antibiorezistente
|
Bacterii patogene |
Antibioticul indicat pentru organismul susceptibil |
Antibiotice alternative daca organismul este rezistent la medicamentul indicat |
|
Staphylococcus aureus |
Cloxacilina sau meticilina |
Vancomicina |
|
Streptococcus pneumoniae |
Penicilina |
Cefotaxima, ceftriaxon sau vancomicina * |
|
Haemophilus influenzae |
Ampicilina |
Cefalosporine de generatia III, eritromicina, TMP/SMX, Cefaclor§, Amoxicilina/clavulanat, Claritromicina |
|
Enterococcus fecalis |
Ampicilina si gentamicina |
Vancomicina |
|
Moraxella catarrhalis |
Ampicilina |
La fel ca si pentru H. influenzae |
|
Mycobacterium tuberculosis |
Izoniazida (in combinatie cu rifamfin si alte medicamente) |
O combinatie de 3-6 medicamente la care organismul este susceptibil |
|
Specii de Shigella |
Ampicilina sau TMP/SMX |
Cefixim, ceftriaxon, cefotaxim, ciprofloxacin¶ |
|
Specii de Salmonella |
Ampicilina |
TMP/SMX, cloramfenicol, ceftriaxon |
|
Pseudomonas aeruginosa |
Ticarcilin si tobramicina |
Ceftazidim, cefoperazon, quinolone¶, imipenem |
|
* Alegerea este determinata de localizarea infectiei si de rezultatele antibiogramei; terapie orale; Trimetoprim/sulfametoxazol (TMP/SMX); §Sau alta cefalosporina orala de a doua sau a treia generatie; A nu se folosi in cazul copiilor |
||
|
Politica de confidentialitate |
| Copyright ©
2025 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |