TRANSMITEREA INFORMATIEI EREDITARE
Informatia
genetica, necesara pentru sinteza diferitelor proteine si
realizarea caracterelor fenotipice, se transmite din generatie in
generatie, in doua etape:
- sinteza unor molecule de ADN
identice cu molecula initiala prin replicare semiconservativa,
care dubleaza cantitatea de ADN;
 - distribuirea completa,
egala si exacta a materialului genetic dublat, prin diviziunea
celulara.
- distribuirea completa,
egala si exacta a materialului genetic dublat, prin diviziunea
celulara.
Ambele procese sunt supuse unor
controale riguroase si se realizeaza de obicei cu mare exactitate,
asigurand astfel stabilitatea proceselor ereditare. Ele pot suferi insa
si erori; daca lipsesc mijloacele eficiente de reparare a acestor
erori, ele pot avea consecinte importante la descendenti, determinand
boli.
1.
REPLICAREA ADN
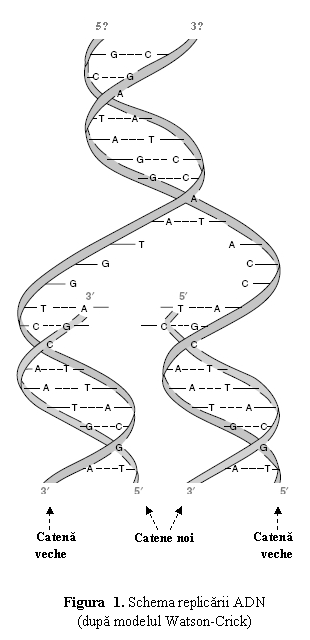
Asa cum au intuit Watson
si Crick (1953), mecanismul replicarii este sugerat de
complementaritatea catenelor polinucleotidice din dublul helix ADN: fiecare catena
serveste drept matrita pentru formarea unei catene noi (fig. 1).
In
esenta, molecula de ADN se despiralizeaza si cele doua
catene se separa prin ruperea legaturilor de hidrogen dintre bazele
complementare (asemanator deschiderii unui fermoar), formand o
structura de forma literei Y, numita furca de replicare. Fiecare catena actioneaza
ca matrita (tipar) pentru aranjarea complementara, in sensul
5'→3' a dezoxiribonucleotidelor activate, care vor fi polimerizate sub
actiunea ADN-polimerazei. Astfel, molecula dublu catenara de ADN
formeaza doua molecule noi, identice atat intre ele cat si cu
molecula parentala; fiecare molecula este formata dintr-o
catena "veche" si o catena nou-sintetizata, procesul de
sinteza al ADN fiind astfel numit replicare
semiconservativa.
Replicarea ADN este mai bine
cunoscuta la procariote, dar mecanismul de baza este similar si
la eucariote. Replicarea ADN la celulele animale este mult mai complexa
datorita dimensiunii foarte mari a genomului, fragmentarii in
cromozomi, asocierii cu proteine in nucleozomi si condensarii. De
aceea, procesul implica numeroase interactiuni intre proteine si
intre acestea si ADN.
1.1. Elemente implicate in replicarea
ADN
(1)
Dezoxiribonucleotide activate (dezoxiribonucleozide
trifosfat - dNTP): dezoxiadenozintrifosfat (dATP), dezoxiguanozintrifosfat
(dGTP), dezoxicitidintrifosfat (dCTP) si dezoxitimidintrifosfat (dTTP). Ruperea
legaturilor fosfat macroergice dintre fosfatii si elibereaza
doua grupuri fosfat si formeaza dezoxiribonucleotide activate.
(2)
Matrita (tipar) este reprezentata de fiecare
catena parentala de ADN.
(3) Enzimele
care catalizeaza sinteza de ADN sunt numite ADN-polimeraze. La om au fost identificate cinci ADN-polimeraze,
notate α, β, γ, δ si ε; replicarea ADN nuclear
este realizata de ADN-polimeraza
δ, enzima majora a replicarii si ADN-polimeraza α;
ADN-polimeraza γ realizeaza replicarea ADN mitocondrial, iar ADN-polimerazele
β si ε sunt implicate in repararea ADN.
(4)
ADN-helicazele catalizeaza desfacerea dublului helix de ADN (folosind
energie rezultata din ruperea legaturilor fosfat macroergice) si
elibereaza monocatenele matrita. Helicazele actioneaza
in puncte specifice numite origini de replicare, asigurand apoi inaintarea
furcii de replicare prin ruperea legaturilor de hidrogen, despiralizarea
si deschiderea moleculei, pentru a deveni accesibila diferitelor
proteine ale replicarii.
(5)
Proteinele de replicare A (RPA), numite si proteine de legare a monocatenelor ADN sau proteine SSBP (single-strand
binding protein) mentin separate cele doua catene desfacute
de helicaze; SSBP se fixeaza la monocatenele ADN si impiedica reimperecherea
bazelor, deci respiralizarea.
(6)
ADN-topoizomerazele I si II Despiralizarea
dublului helix fara rotatia acestuia creaza in aval de
furca de replicare niste superhelixuri (asemanator separarii
bruste a firelor rasucite dintr-o franghie); topoizomerazele rup
legaturile fosfodiester la nivelul uneia (topizomeraza I) sau a ambelor catene
(topizomeraza II), produc rotatia libera a capetelor helixului
si apoi resudarea lor; in acest fel topoizomerazele detensioneaza
(relaxeaza) molecula de ADN.
(7) Primaza este o ARN-polimeraza
specifica implicata in sinteza primerului si replicarea catenei
"intarziate". ADN-polimerazele nu pot initia replicarea, o pot doar
continua. Astfel, sub actiunea primazei se sintetizeaza primerul, un fragment scurt de ARN (10-12 nucleotide), complementar cu catena matrita; ADN-polimeraza
α adauga, la capatul OH 3' al primerului, circa 30 de
dexoxiribonucleotide. Primaza si ADN-polimeraza α sunt apoi
indepartate, sinteza lantului continuand sub actiunea ADN-polimerazei
δ.
(8)
Proteinele de replicare C
(RPC) se leaga la jonctiunea dintre primer si
matrita, stabilizand interactiunea ADN-polimerazei cu
matrita ADN.
(9)
Antigenul nuclear de proliferare celulara (PCNA - Proliferating Cell Nuclear Antigen) se
leaga in imediata vecinatate a proteinelor C; ambele controleaza
pornirea sau oprirea replicarii.
(10)
Ribonucleaza H1 (RNaza H1) indeparteaza primerul ARN,
folosit pentru initiera replicarii; lacuna ramasa este
completata de ADN-polimeraza δ, iar sudarea capetelor este
realizata de catre o ADN-ligaza,
refacand continuitatea catenei.
1.2. Mecanismul molecular al
replicarii ADN
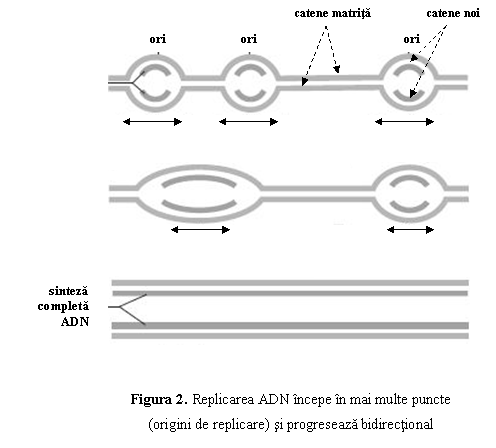
Procesul
de replicare incepe in puncte specifice numite origini de replicare (ori) si, de aici, progreseaza in ambele directii, pana ce ADN
este complet duplicat (fig. 2). La nivelul originilor de replicare, pe ADN se
fixeaza enzimele replicarii: topoizomerazele, helicazele, primazele,
ADN-polimerazele.
Dublul
helix este despiralat sub actiunea topoizomerazelor,
iar catenele sunt separate temporar de catre helicaze, formand furca de
replicare, o structura in forma de Y. Pe fiecare catena
separata se fixeaza proteinele SSB (RPA), care impiedica
reimperecherea bazelor complementare si refacerea spontana a dublului
helix.
Pe cele
doua catene matrita, ADN-polimeraza δ ataseaza
secvential si complementar dezoxiribonucleotidele activate.
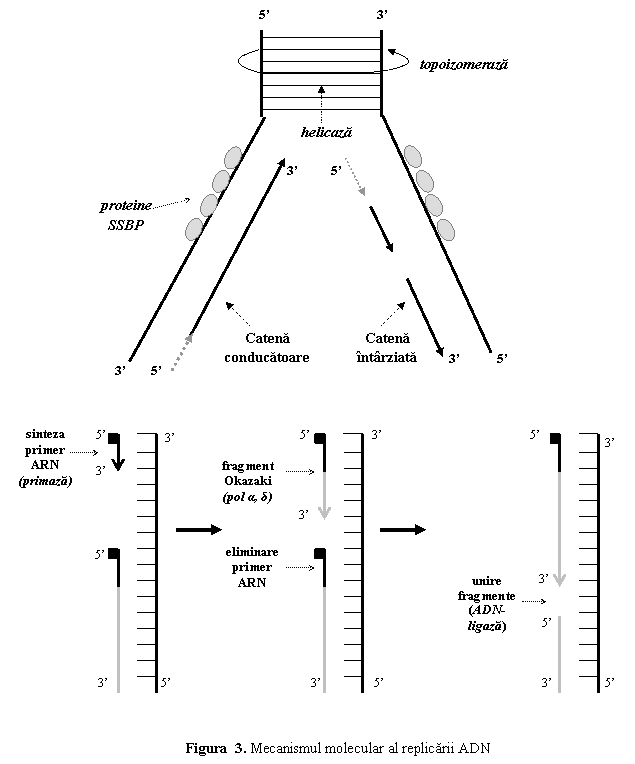
Polimerizarea
nucleotidelor se face diferit pe cele doua catene, care sunt antiparalele.
ADN-polimeraza nu poate incepe sinteza prin legarea a doua nucleotide; ea
poate doar sa adauge nucleotide la capatul 3'OH al unui lant
preexistent (ADN-polimeraza poate numai sa alungeasca catena in curs
de sinteza); de aceea necesita utilizarea unui primer ARN, sintetizat
sub actiunea primazei. Sinteza se poate efectua numai in sensul 5'-3'. Astfel
numai una din cele doua catene, denumita catena conducatoare sau directoare (leading strand)
(catena care are ca matrita catena 5'-3') poate fi sintetizata in mod continuu si in sensul
deplasarii furcii de replicare.
Cealalta
catena - catena intarziata (lagging strand) (catena care are ca matrita catena 3'-5') - este sintetizata discontinuu
si mai lent, sub forma de secvente scurte (100-1000 nucleotide)
denumite fragmente Okazaki; sinteza
fiecarui fragment are loc in sens opus celui in care se deplaseaza
furca de replicare.
Primerul este eliminat prin hidroliza si
inlocuit cu secvente ADN, sub actiunea ADN-polimerazei δ, iar
fragmentele sunt unite de catre o ADN-ligaza. Primerul este necesar
numai la inceputul replicarii pentru catena conducatoare, iar pentru
catena intarziata la sinteza fiecarui fragment Okazaki (fig. 3).
1.3. Particularitatile
replicarii ADN la eucariote
Datorita
dimensiunii mari a genomului si asocierii cu proteine, viteza de replicare
este mai redusa - circa 50 nucleotide/secunda (fata de 500
nucleotide/secunda la procariote).
Replicarea incepe in mai multe
puncte de origine (secvente specifice de ADN) care corespund
unitatilor de replicare numite repliconi.
Replicarea progreseaza bidirectional pentru fiecare replicon si,
dupa ce se termina, repliconii fuzioneaza treptat pana ce
intregul cromozom este duplicat.
(2) Comportamentul proteinelor
nucleozomale in timpul replicarii inca nu este cunoscut cu precizie;
se stie insa ca, odata cu siteza ADN, in nucleu are loc
si o intensa sinteza de histone, necesare formarii de noi
nucleozomi.
(3)
Replicarea are loc in faza S a
ciclului celular si dureaza, in celulele umane, circa 8 ore (la un
ciclu celular de 24 de ore). Deoarece acest timp este scurt pentru replicarea
celor peste 6 miliarde de pb din genomul uman, uneori pot apare erori.
Relicarea este reglata de o serie de proteine:
. diferite cicline asigura
trecerea din faza G1 in faza S, precum si progresia prin faza
S;
. CDK4 este
o protein-kinaza activatoare
. anumiti factori de transcriptie
activeaza punctele de origine, iar altii stimuleaza expresia
genelor necesare intrarii in faza S;
. Inhibitorii kinazelor dependente
de cicline - CKI - blocheaza intrarea si progresia prin faza S
(prin inhibarea complexelor CDK-cicline si PCNA) atunci cand apar leziuni
ale ADN sau cand un tesut a incheiat diferentierea.
Replicarea celor 20-80 de repliconi nu
este sincrona si se realizeaza intr-o anumita
ordine, in functie de structura cromatinei: la inceputul fazei S
este replicata eucromatina, iar la sfarsitul fazei S este
replicata heterocromatina.
In mod normal, intregul genom este
replicat in faza S o singura data.
(4)
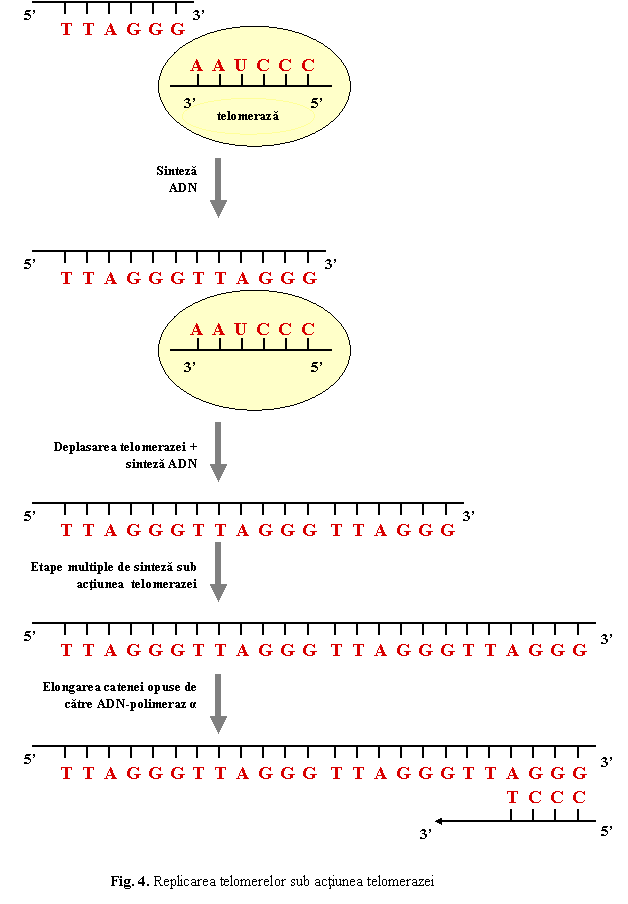
Replicarea telomerelor
Telomerele sunt alcatuite din
secvente hexamerice repetitive TTAGGG dispuse in tandem pe o lungime de
5-20 kb. Un fenomen caracteristic telomerelor este pierderea de secvente ADN la fiecare replicare, deoarece, la
capatul 5' al catenelor nou sintetizate replicarea este incompleta, pierzandu-se la fiecare ciclu
celular intre 25 si 200 pb. Aceasta duce la scurtarea progresiva a telomerelor si, dupa un
numar de replicari (maximum
80 la celulele somatice, cand este atinsa limita critica de circa 1,5
kb), la oprirea replicarii si
a diviziunii.
La
nivelul celulelor germinale, in succesiunea generatiilor, lungimea
telomerelor ramane nemodificata datorita actiunii enzimei telomeraza.
Telomeraza este o reverstranscriptaza speciala care contine o
secventa scurta de ARN, complemetara secventelor
repetitive telomerice; telomeraza adauga secvente TTAGGG la
capatul 3', fara a necesita o matrita (fig. 4.).
In
majoritatea celulelor somatice, expresia genei telomerazei este inhibata
inca din stadiul embrio-fetal. Disparitia activitatii
telomerazei dupa nastere determina scurtarea progresiva a
telomerelor dupa fiecare diviziune, pe masura inaintarii in
varsta. Cand este atinsa o lungime critica, diviziunea se
opreste si incepe senescenta. Pierderea telomerelor produce, de
asemenea, instabilitate genomica, urmata adesea de rearanjamente
cromozomiale.
Telomeraza este reactivata in celulele
canceroase (vezi Cap. Boala canceroasa).
Procesul
de replicare este realizat de catre ADN-polimeraze cu mare fidelitate:
rata erorilor de imperechere (mismatch)
este de circa 1/10
ADN-polimeraza are, in primul rand,
capacitatea de a pozitiona corect
bazele complementare (probabil pe baza conformatiei tridimensionale). In
al doilea rand, ADN-polimerazele δ si ε au activitate de corectare (proofreading), inlaturand nucleotidele gresit incorporate
printr-o actiune 3'-5'- exonucleazica.
Putinele erori de imperechere
care apar in prezenta mecanismelor mentionate sunt reparate.
Deplasarea telomerazei + sinteza ADN
|
Etape multiple de sinteza sub actiunea telomerazei
|
Elongarea catenei opuse de catre ADN-polimeraz α
|
Fig. 4. Replicarea telomerelor sub actiunea
telomerazei
|