
GENETICA DROJDIILOR
Introducere
Drojdiile reprezinta un grup de microorganisme eucariote, a caror denumire defineste grupul din punct de vedere morfologic, structural si genetic. Studiile recente de filogenie moleculara au demonstrat insa ca drojdiile nu reprezinta un grup separat, fiind formele unicelulare ale unor fungi filamentosi.
In prezent, se pastreaza inca gruparea drojdiilor in functie de prezenta/absenta si mecanismele procesului de sexualitate, in : ascomicete, basidiomicete si deuteromicete (fungi imperfecti). Cu toate acestea, analizele moleculare indica faptul ca ascomicetele de tipul Saccharomyces si Pichia reprezinta formele teleomorfe (forme ce prezinta meioza si sporulare si, implicit, sexualitate) ale fungilor imperfecti de tipul Candida, considerati ca forme anamorfe (forme care se divid vegetativ prin mitoza si nu prezinta sexualitate). Dintre ascomicete, drojdia care a reprezentat modelul experimental pentru majoritatea studiilor biologice este Sacchromyces cerevisiae, microorganism care constituie si obiectul principal al problemelor teoretice si tehnicilor descrise in acest manual.
Organizarea unicelulara care presupune crestere si multiplicare rapida, propagarea clonala, posibilitatea de marcare selectiva si analiza genetica, precum si structura eucariota, a permis descifrarea unor aspecte legate de ciclul celular, structura si replicarea ADN la eucariote, structura genei eucariote, precum si descifrarea mecanismelor de reglaj genetic, de splicing, de mutatie si recombinare genetica. Astfel, o serie de aspecte legate de genetica si biochimia celulei eucariote, explicate initial pornind de la sisteme procariote, au putut fi intelese mai acurat prin studiile realizate pe S. cerevisiae.
1. Ciclul de viata la Saccharomyces cerevisiae
Ciclul de viata la S. cerevisiae reprezinta, pe de o parte, un proces de proliferare celulara prin care o celula de un anumit tip de imperechere (a sau prin diviziune mitotica da nastere la doua celule in esenta identice, iar pe de alta parte, procesul prin care se modifica gradul de ploidie, existand atat forme diploide cat si haploide stabile. Haploizii se imperecheaza pentru a forma diploizi ce se pot divide meiotic in cursul procesului de sporulare, formand haploizi de tip de imperechere a si Aceste procese de imperechere si meioza sunt procese biologice fundamentale care apar si la organismele pluricelulare. Totusi, ciclul de viata la S. cerevisiae prezinta un aspect particular in sensul ca, celulele haploide, pe langa proliferare, imperechere si meioza, pot suferi rearanjamente cromozomiale ce stau la baza interconversiei tipului celular de imperechere.
Tulpinile de S. cerevisiae apartin la doua tipuri de fertilizare: tulpini homotalice care prezinta un proces de autofertilizare formand diploizi pornind de la o cultura monosporala si tulpini heterotalice care prezinta fertilizare incrucisata si formeaza diploizi prin imperecherea celulelor a si de tip opus de imperechere (Fig.1.).
In cazul tulpinilor heterotalice pornind de la o celula diploida heterozigota (a/ ) pentru locusul tipului de imperechere (MAT = mating type) in urma procesului de meioza si sporulare (pe medii specifice de sporulare sarace in nutrienti) se formeaza o asca cu 4 ascospori (2a/2 Fiecare celula haploida a sau cultivata separat pe medii vegetative (YPG) formeaza in urma diviziunii mitotice prin inmugurire clone haploide stabile a si . Daca aceste celule sunt cultivate impreuna pe mediu vegetativ, in urma recunoasterii celulare pe baza feromonilor a si , fuzioneaza realizandu-se un proces de citogamie si kariogamie, rezultand celule diploide ce pot forma de asemenea, prin diviziune mitotica, clone diploide stabile.
La tulpinile homotalice, pornind de la un singur ascospor haploid a sau , in urma diviziunii mitotice rezulta datorita interconversiei tipului de imperechere, o cultura mixta a si , care genereaza, in final, numai diploizi stabili.
Intelegerea interconversiei tipului de imperechere prespune intelegerea structurii genetice a locusului MAT si a celor 2 casete silentioase plasate la stanga (HML = homotalic left) si la dreapta (HMR = homotalic right) fata de acesta si care poarta informatie suplimentara a si . Cei 3 loci sunt dispusi pe cromozomul III iar in mod obisnuit nu se exprima decat locusul MAT, casetele fiind represate prin cuplarea unor proteine specifice codificate de genele SIR (silent information regulator). Structura locusului MAT si a celor 2 casete este, in esenta, identica, in cazul segmentelor W, X, Z, fiind variabil numai segmentul Y de aproximativ 600 pb care codifica fie pentru factorul a (alela a), fie pentru factorul (alela ). (Fig. 2.).
O alta gena importanta in procesul de interconversie este gena HO care codifica pentru endonucleza Ho (Y/Z) care recunoste o secventa specifica de baze prezenta la jonctiunea segmentelor Y/Z producand incizii dublucatenare numai la nivelul locusului MAT, casetele fiind protejate de proteinele Sir prin condensare. Gena HO (dominanta) este prezenta la tulpinile homotalice, in timp ce la tulpinile heterotalice este prezenta alela recesiva ho, fapt care nu permite interconversia tipului de imperechere la aceste tulpini.
Interconversia tipului de imperechere se realizeaza in mai multe etape care presupun:
- realizarea unei incizii dublucatenare la jonctiunea Y/Z;
- degradarea segmentului Y din locusul MAT;
- formarea unor capete libere 3' care reprezinta primeri pentru o polimeraza reparatorie care copiaza informatia de pe segementul Y apartinand casetei cu informatie corespunzatoare tipului opus de imperechere;
- sinteza reparatorie determina, in final, inlocuirea unui segment Y, de exemplu Ya, cu segmentul complementar Y . Fiecare diviziune mitotica presupune parcurgerea procesului de interconversie, fapt care determina formarea culturii mixte de haploizi caracteristica tulpinilor homotalice.
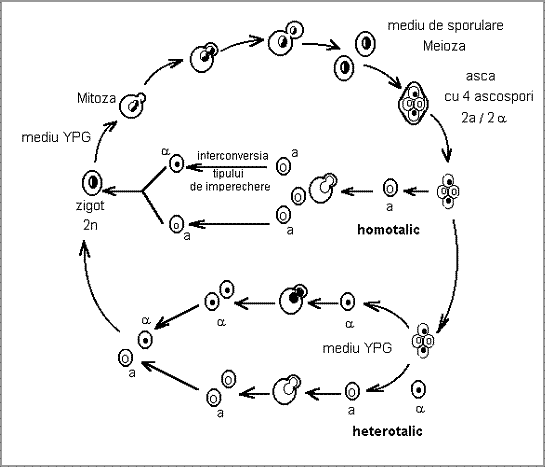
Fig.1. Ciclul de viata la Saccharomyces cerevisiae
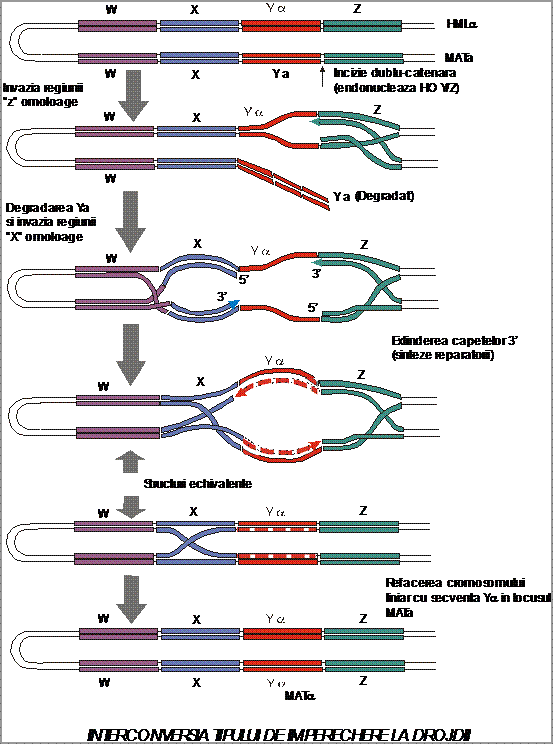
Fig. 2. Interconversia tipului de imperechere la Saccharomyces cerevisiae
Definirea complexitatii ciclului celular la drojdii a pus problema identificarii genelor implicate in controlul diferitelor ciclicuri. Aceste gene au putut fi caracterizate prin analiza unor mutante specifice - asa numitele mutante cdc (cell division cycle). Acestea sunt mutante conditionale care la temperaturi permisive exprima fenotipul de tip salbatic. Studiul mutantelor cdc a permis stabilirea faptului ca celulele haploide pot intra in mitoza sau meioza in functie de genele cdc activate. Astfel, in cazul in care inaintea punctului START, sub actiunea factorilor de imperechere a sau , sunt activate genele cdc36 si cdc39, celulele haploide sunt blocate in G1, se imperecheaza (a x ) formandu-se diploizi care intra intr-un proces de sporulare si meioza (Fig. 3.).

Fig. 3. Rolul genelor cdc in controlul ciclului celular la Saccharomyces cerevisiae
Gena cruciala care determina trecerea peste punctul START si desfasurarea mitozei, este gena cdc28, al carui produs este o proteinkinaza, fosforilarea proteinelor fiind esentiala in desfasurarea diviziunii celulare. Functiile cdc28 sunt activate de acumularea ciclinelor (Cln 1, 2, 3) la sfarsitul fazei G1, fiind facilitata trecerea in faza S. Trecerea din S in G2/M este controlata tot de gena cdc28, activarea fiind realizata insa, de ciclinele de tip B - Clb 1, 2. (Fig ).
Alte gene cdc care se exprima in etapa de trecere de la G1 la S sunt: cdc7 (proteina codificata este o proteinkinaza implicata in fosforilarea proteinelor ce participa in replicare), cdc17 (produsul ei este polimeraza ), cdc2 (polimeraza ), cdc21 (codifica o timidilatsintetaza), cdc8 (timidinkinaza) si cdc9 (ADN ligaza) (Fig. ).
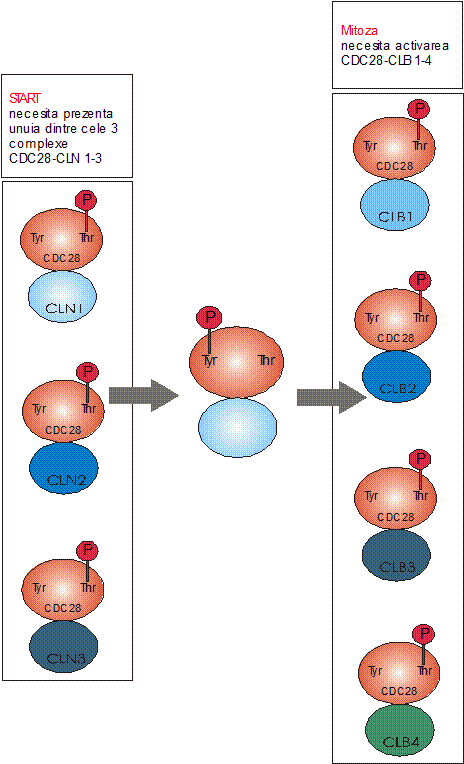
Fig. Activarea Cdc28 de catre diferite cicline determina trecerea de START sau intrarea in mitoza la Saccharomyces cerevisiae
Cap. 2. Organizarea materialului genetic la Saccharomyces cerevisiae
2. 1. Structura genomului nuclear la Saccharomyces cerevisiae
Genomul nuclear la drojdii are o structura cromatinica specifica eucariotelor cu o compozitie histonica normala in miezul nucleosomal, cu o dimensiune per genom haploid de 1.3 x 107 (14 X 103 kpb). Genomul este structurat in 16 cromosomi cu dimensiuni cuprinse intre 245 kpb (cromozomul I) si aproximativ 2500 kpb (cromozomul XII), cuprinzand 6500 de gene.
In afara genelor structurale ce codifica pentru proteine, genomul drojdiilor contine 140 gene pentru ARNribozomal (ARNr) prezente intr-un numar variabil de copii (100 - 200) pe cromozomul XII, circa 40 de gene repartizate pe diferiti cromosomi care codifica small nuclear ARN si circa 400 de gene pentru ARNtransfer (ARNt). Dintre toate aceste gene numai 4% prezinta introni, acestea fiind, gena pentru actina (care contine 1 intron), gena MAT 1 si 40 gene pentru ARNt.
Principalele substructuri cromosomale evidentiate si studiate la drojdii sunt secventele ARS (autonomously replicating sequence) echivalente cu originile de relicare (ORI), secventele CEN (centromerice) si TEL (telomerice).
Secventele ARS
Replicarea ADN incepe de la nivelul originilor de replicare ARS, prima secventa identificata fiind ARS 1 (Bel si Stillman ,1979). La nivelul genomului drojdiilor exista aproximativ 400 ARS, care se succed la circa 35 kpb, replicarea ADN incepand la momente diferite la nivelul unor clustere formate din 20 - 80 secvente ARS care formeaza unitati replicative. Secventa ARS1, cu o dimensiune de 50 pb, este structurata in mai multe unitati: A si B, cea din urma fiind structurata, la randul ei, in 3 subunitati: B11, B21 si B31. Secventa esentiala este ACS (ARS consensus sequence) de 11 pb, la nivelul careia se leaga complexul proteic ORC (origin recognation complex) cu rolul de initiere a replicarii. Cele 3 subunitati B sunt secvente aditionale ale ACS, partial identice: B11 - este secventa suplimentara de legare a ORC, B21 - are rol structural si B31 este o secventa de legare pentru factorul de transcriere OBF/ABF (origin/ARS binding factor) care stimuleaza replicarea prin domeniul activator transcriptional (Fig. 5.).
![]()
Fig. 5. Structura secventei ARS1 de la Saccharomyces cerevisiae
Secventele CEN
Prima secventa centromerica descrisa a fost CEN3 de la nivelul cromosomului III (Clarke si Carbon, 1980). La S. cerevisiae centromerii sunt formati din 3 secvente: CDE I (9 pb) - secventa de baze inalt conservata, plasata in partea stanga, CDE II (78 - 89 pb) - cu un continut de peste 90% AT, prezenta la toti centromerii, cu o constitutie asemanatoare cu ADN satelit de la eucariotele superioare si CDE III (11pb) - secventa inalt conservata (TGATTTCGGAA), secventa consens de legare a factorului CBF (centromer binding protein). Factorul CBF este un complex proteic de aproximativ 6 proteine esentiale in structurarea kinetocorului.
Actualmente, cel mai bine caracterizate sunt secventele CEN3, CEN4 si CEN11 (Fig.
CDE I CDE II CDE III
CEN3 ATAAGTGACATGAT - 88 pb (93% AT) - TGATTTCCGAA
CEN4 ATAAGTGACATGAT - 89 pb (94% AT) - TGATTTCCGAA
CEN11 ATAAGTGACATGAT - 82 pb (83% AT) - TGATTTCCGAA
Secventele TEL
Secventele TEL si la drojdii sunt secvente scurte repetate in tandem, plasate la capatul cromosomilor, cu o structura non-cromatidica, denumita telosom. Fiecare secventa TEL este structurata in:
- unitate repetitiva - are secventa C1-3A/TG1-3;
- aditional acestor repetari la capetele extreme ale cromosomilor, telomerele prezinta si ADN mediu repetitiv reprezentat de elemente ADN asociate (telomere associated - TA): Y' si X. Y' este un element inalt conservat care poate avea 2 dimensiuni diferite - 6.7 kpb si 5,2 kpb si poate exista intr-un numar variabil de copii (0 - 4 copii/telomer). X cuprinde mai multe secvente repetitive mici de 35 - 560 pb, dimensiunea elementului X variind intre 0.3 kpb si 3.75 kpb;
- intre cele 2 elemente X si Y' exista un numar mic de secvente repetitive TG1 -3 (Fig.
![]()
Fig. 7. Structura regiunii TEL la Saccharomyces cerevisiae
3. Plasmida 2 m
Denumita astfel dupa lungimea conturului, determinat prin studii de microscopie electronica, plasmida 2 m este formata dintr-o molecula de ADN dublu catenar circular covalent inchis, avand o dimensiune de 6.3 Kpb (2 MD). Aceasta plasmida este prezenta la majoritatea tulpinilor de S. cerevisiae in 60 - 100 copii / celula.
In studiile initiale, plasmida 2 m a fost considerata un element genetic citoplasmatic, dar, analizele ulterioare au demonstrat, pe de o parte, localizarea sa nucleara, iar pe de alta parte, structura sa nucleozomala.
Trasatura cea mai importanta a plasmidei 2 mm este reprezentata de 2 secvente invers repetate (IR) de 599 pb fiecare, care separa molecula in doua regiuni cu secventa unica, de 2774 pb, si respectiv, 2346 pb (Fig. 8). Cele 2 regiunile repetate invers permit recombinari intramoleculare si, ca o consecinta, plasmida se poate gasi in doua forme echivalente, A si B, care difera prin orientarea relativa a celor 2 secvente unice.
In urma recombinarilor intermoleculare la nivelul regiunilor repetate invers pot sa apara multimeri de 4 m si 6 m. Trecerea de la o forma moleculara la alta (proces denumit flipping ) nu se realizeaza printr-un proces care sa implice si o transpozitie a plasmidei pe ADN cromosomial, deoarece nu s-au identificat secvente corespunzatoare plasmidei 2 mm in ADN cromosomial.
Plasmida 2 mm prezinta 4 gene:
a gena FLP (flipping) - codifica o proteina ce catalizeaza recombinarea situs-specifica la nivelul secventelor repetate invers;
a genele REP 1 si REP 2 (replication) - codifica proteine ce controleaza, pe de o parte, stabilitatea si segregarea plasmidei 2 m, iar pe de alta parte, exprimarea genelor plasmidiale (4);
a gena RAF 1 - codifica o proteina implicata tot in reglarea exprimarii genice.
In structura plasmidei 2 m au mai fost identificate urmatoarele secvente functionale:
- secventa ARS (autonomously replicating sequence) reprezinta originea replicarii plasmidei si este plasata la nivelul jonctiunii dintre regiunea unica mica si una din regiunile repetate invers, astfel incat cuprinde o parte din regiunea IR si 100 bp din regiunea unica mica ; regiunea FRT (FLP recognition target - tinta de recunoastere pentru FLP) la nivelul careia actioneaza recombinaza FLP; regiunea FRT este situata in interiorul uneia din cele 2 IR ; secventa STB (stability) necesara segregarii stabile a plasmidei in mitoza, situs de actiune pentru produsii genelor REP 1 si REP 2.
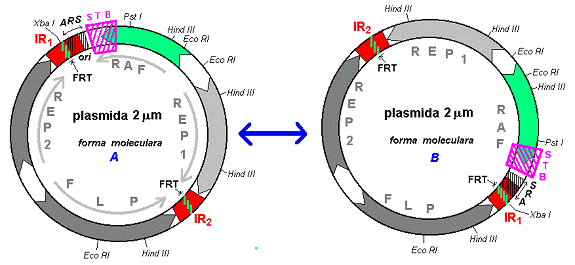
Fig. 8. Structura si formele moleculare ale plasmidei 2 m
Materialul genetic extranuclear
1. Genomul mitocondrial la Saccharomyces cerevisiae
Mitocondria este un organit complex, specializat in respiratia celulara si fosforilarea oxidativa, care prezinta un sistem genetic propriu si, implicit, mecanisme specifice de sinteza proteica. Spre deosebire de celelalte organisme eucariote la care procesele de fosforilare oxidativa sunt catalizate de 5 complexe enzimatice, la drojdii nu exista decat 4 complexe enzimatice, dintre care 3 complexe respiratorii si un complex ATP-azic (ATP-sintetaza), lipsind complexul reprezentat de NADH-CoQ-reductaza :
I - succinat-CoQ-reductaza: oxideaza succinatul, reducandu-l la fumarat si reduce CoQ
II - CoQH2-citocrom c-reductaza: oxideaza CoQ si reduce citocromul c
III - citocrom-oxidaza: transfera electronul la oxigen reducand citocromul c si rezultatul este formarea apei
IV - ATP-aza mitocondriala, care intervine in etapele fianle ale fosforilarii oxidative (in stare cuplata functioneaza ca o ATP-sintetaza sau kinaza).
Biogeneza mitocondriei reprezinta un proces complex, care necesita interactiunea a doua genomuri - nuclear si mitocondrial.
La drojdii ADN-ul mitocondrial (ADNmt) este reprezentat de o molecula de ADN dublu catenar circular inchis, asemanator cromosomului bacterian, avand o dimensiune de aproximativ 25 kbp (50 Md). Principalele caracteristici ale ADN-ului mitocondrial de drojdii sunt urmatoarele : (1) este necomplexat cu histone, ci cu proteine histone-like; (2) are un procent mare de AT (80%); (3) are multe gene mozaicate, spre deosebire de genele nucleare de la drojdii care sunt, in general, nemozaicate (exceptii: gena pentru actina, ADNr); (4) exista unele diferente intre codul genetic nuclear si cel mitocondrial (Tab. 1).
Tab. 1. Diferente intre codul genetic universal si cel mitocondrial
|
CODON |
Mesaj in codul universal |
Mesaj in codul mitocondrial |
|||
|
mamifere |
Drosophila |
drojdii |
plante |
||
|
UGA |
Stop |
Trp |
Trp |
Trp |
Stop |
|
AUA |
Ile |
Met |
Met |
Met |
Ile |
|
CUA |
Leu |
Leu |
Leu |
Thr |
Leu |
|
AGA, AGG |
Arg |
Stop |
Ser |
Arg |
Arg |
ADNmt la drojdii cuprinde urmatoarele gene (Fig. 9.): circa 24 specii moleculare de ARN (aceste gene sunt grupate intr-un cluster); 2 specii moleculare de ARNr: 15S in subunitatea de 30S, 21S in subunitatea de 50S (ARNr de 5S este absent); gena pentru ARN 9S (ARN-9S va complexa o proteina, formand o ribozima denumita RNaza P care participa la maturarea ARNt, avand ca situs de actiune capetele 5 ); gene pentru subunitati ale citocrom-oxidazei: CO I, II si III; gena pentru apoproteina citocromului b (partea complexului b-c1, complexul III); genele pentru 3 subunitati ale complexului de sinteza ATP (subunitatile 6, 8 si 9 ale ATPazei mitocondriale); proteine ce interactioneaza cu acizii nucleici. Aceste gene sunt separate (ADN spacer) prin regiuni bogate in A+T.
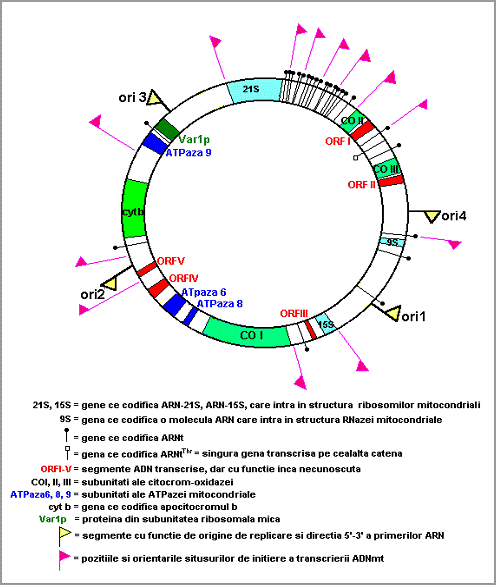
Fig. 9. Harta genomului mitocondrial la Saccharomyces cerevisiae
|
Componenta structurala mitocondriala |
Polipeptide |
|
MATRIX |
carbamil-fosfat-sintetaza |
|
superoxid-dismutaza |
|
|
ADN polimeraza |
|
|
ARN polimeraza |
|
|
proteine ribozomale |
|
|
ornitin-trans-carbamilaza |
|
|
enzimele ciclului acidului citric |
|
|
MEMBRANA INTERN~ |
citocromul c |
|
subunitatea V a complexului citocromului b |
|
|
subunit ale Fi-ATPazei |
|
|
proteolipidul din Fo-ATPaza |
|
|
subunit IV, V, VI, VII ale citocrom-oxidazei |
|
|
SPATIUL INTERMEMBRANAR |
citocromul c |
|
citocrom-peroxidaza c |
|
|
citocromul b2 |
|
|
MEMBRANA EXTERN~ |
porina |
Mutantele petite apar la Saccharomyces cerevisiae corelat cu modificari ale ADN-ului mitocondrial, determinand o serie de deficiente respiratorii; la multiplicare in anaerobioza, mutantele petite formeaza colonii mici ("petite").
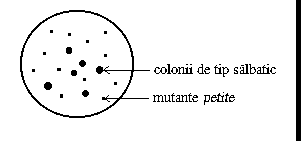
Fig. 10. Aspectul coloniilor petite
Mutantele petite au fost obtinute atat cu mutageni fizici (de exemplu, radiatiile ultraviolete), cat si cu mutageni chimici (nitrosoguanidina). Genomul mitocondrial a fost notat factor r si, ca urmare, tulpinile de tip salbatic sunt r
Agentii mutageni specifici pentru ADN prokariot (de exemplu, acriflavina, bromura de etidium) au actiune primara asupra mitocondriilor, determinand 2 tipuri de mutatii: ro - petite neutre ; r - petite supresive; ro - apar in urma deletiei complete a ADN mitocondrial.
Mutantele petite supresive apar in urma unor deletii partiale ce sunt urmate de duplicatii.
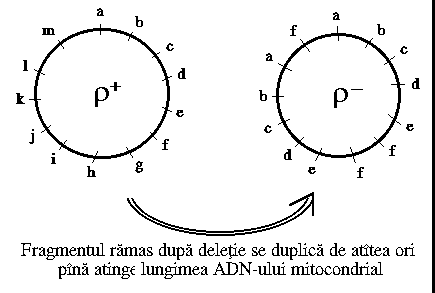
Fig. 11. Structura genomului mitocondrial r si r
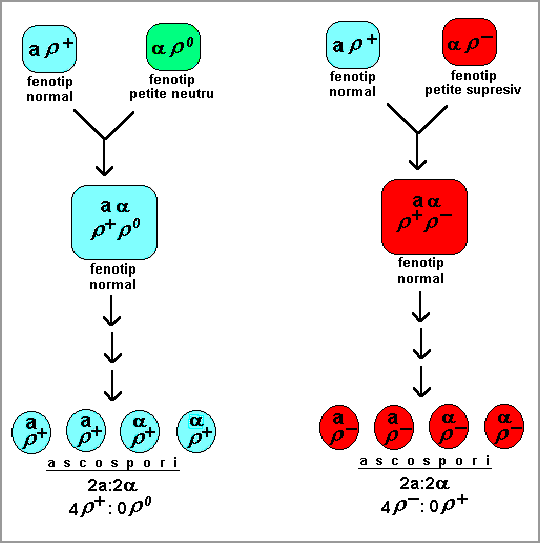
Fig. 12. Schema segregarii nemendeliene a genelor mitocondriale
in cazul diploizilor si
2. Sistemul killer la Saccharomyces cerevisiae
Majoritatea tulpinilor de Saccharomyces cerevisiae au in citoplasma una sau mai multe specii moleculare de virusuri ARN dublu catenar (ARN d.c.). Primele asemnea specii moleculare de ARN d.c. viral descrise sunt cele responsabile de fenotipul killer la drojdii. Aceste molecule prezinta foarte multe similaritati cu virusurile ARN d.c. de la organismele eucariote superioare. Ca urmare, virusurile de la Saccharomyces reprezinta un model pentru descifrarea mecanismelor centrale de replicare virala, de transcriere ARN-dependenta, de impachetare, de frame-shift ribosomal, precum si relatiile dintre functiile virale si cele ale gazdei parazitate.
Virusurile ARN d.c. de la drojdii includ 3 familii de genomuri ARN d.c.: L-A, L-BC si M. L-BC se intalneste mai rar, are aceeasi dimensiune ca si L-A, dar are secventa diferita de nucleotide. Moleculele L-BC sunt localizate in particule virale intracelulare cu proteine capsidale diferite de cele ale capsidelor L-A.
Din aceste motive se considera ca citoplasma tulpinilor killer de la S.cerevisiae cuprinde 2 tipuri de particule: L si M. Particulele M sunt notate VSc-M si codifica toxina si factorul de imunitate, iar particulele L sunt notate VSc-L, codifica capsidele M si L, precum si polimeraze necesare replicarii ambelor genomuri, si au rol de helper in procesul de infectie.
Particulele killer de la S.cerevisiae reprezinta molecule de ARN d.c. incapsidate in particule virus-like (cu un diametru de 40 nm) si sunt responsabile de sinteza toxinei killer, substanta letala pentru tulpinile de drojdii non-killer.
Sistemul killer a fost descoperit pentru prima oara la Saccharomyces in 1963, ulterior fiind descris si la alte genuri si specii de drojdii. In prezent se cunosc 11 tipuri de toxine killer biochimic diferite, produse de diverse genuri de drojdii.
Proteina de fuziune gag-pol are 3 domenii functionale (Fig. 13):
domeniu capsidal (intra in structura capsidei, alaturi de proteina majora a capsidei)
domeniu de atasare a ARN m.c. (+)
domeniu cu functie de ARN-polimeraza
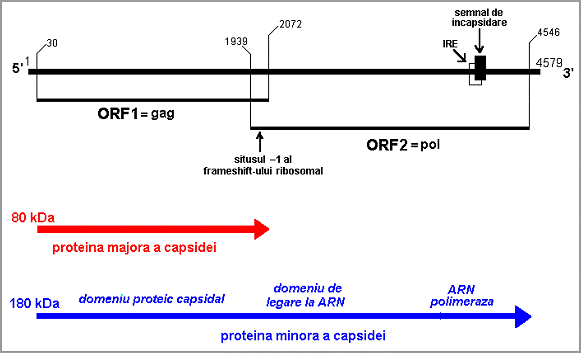
Fig. 13. Structura genelor gag-pol din genomul L
Daca particulele L pot contine fie o molecula ARN d.c.-L, fie una m.c.-L (+), particulele M pot contine una sau doua molecule ARN d.c.; in ambele cazuri, particulele M au aceeasi dimensiune si compozitie proteica, care este cvasi-identica cu particulele L.
Transcrierea genomurilor M
ARN d.c. al particulelor M codifica pentru toxina si pentru imunitate la aceasta.
Activitatea killer a tulpinilor de S.cerevisiae apartine la 3 tipuri - K1, K2 si K3 - codificate de genomurile M1, M2 si, respectiv, M3: K1 - tulpinile de laborator; K2 si K3 - tulpinile de vin si de bere. Tulpinile K1 si K2 sunt sensibile la toxina fiecaruia, in timp ce K2 si K3 sunt cros-imune. Lipsa totala a particulelor killer a fost notata drept K0. Rezistenta este notata cu R.
Genomul M1 are un singur ORF ce se extinde de la codonul AUG (aflat in pozitia 14) pana la codonul UAG (aflat la pozitia 963).
L M
(100 copii/cel) (10-12 copii/cel)
K1 2.44 x 106 Da 1.28 x 106 Da
K2 2.54 x 106 Da 1.00 x 106 Da
Atat particulele M, cat si cele L, nu au caracter infectios si nu prezinta ciclu litic. Ele sunt virusuri latente (cu o serie de caractere de plasmide) si pot fi transmise prin imperechere sexuala sau prin fuziune de protoplasti.
ARN d.c.-M cuprinde un fragment de 1kb separat prin 200 bp (regiune bogata in AU) de un fragment de 0.6 kbp. Fragmentul de 1 kbp poarta informatia genetica pentru precursorul toxinei de 34000 Da.
Precursorul toxinei este format din 3 subunitati, si (din care si vor forma toxina, iar va forma factorul de imunitate). Secventa hidrofila leading , cu afinitate mare pentru reticulul endoplasmic, permite penetrarea precursorului in aceasta structura citoplasmatica unde au loc modificari post-translationale de tip glicozilare. In urma maturarii precursorului, toxina va avea o greutate moleculara de 43000 Da (Fig. 13). In timpul transportului intracitoplasmatic, precursorul este fragmentat de proteinaze specifice care separa secventa leader si subunitatea . Proteina killer activa este, deci, formata din subunitatile si unite prin legaturi disulfidice.
Datorita fuziunii veziculelor de transport cu membrana plasmatica, proteina killer si factorul proteic de imunitate sunt eliberate in afara celulei. Factorul proteic de imunitate ramane legat de suprafata celulara, probabil la nivelul unui receptor specific.
Modul de actiune al proteinei killer
Dupa o perioada de lag, proteina killer se leaga de un receptor din peretele celular - 1,6- -D-glucanul - fara consum de energie. Al doilea receptor este plasat probabil in interior, pe membrana plasmatica. Spre interiorul celulei sensibile este transferata doar subunitatea , procesul realizandu-se cu consum de energie.
Efectul letal al toxinei pare sa se datoreze formarii unor pori in membrana plasmatica. Ca urmare a cresterii permeabilitatii membranei, are loc o eliberare a ionilor de K+ si a moleculelor de ATP in mediul extracelular. Aceste procese sunt urmate de o descrestere a pH-ului intracelular si de inhibarea proceselor metabolice, ducand in final la moartea celulara.
Sistemului killer este de o deosebita importanta in procesele biotehnologice, fiind implicat in : (1) protectia procesului de fermentatie la tulpini de drojdii de vin ; (2) procesul de selectie a transformantilor rezultati prin fuziunea de protoplasti ; (3) biotiparea unor tulpini de drojdii si de bacterii.
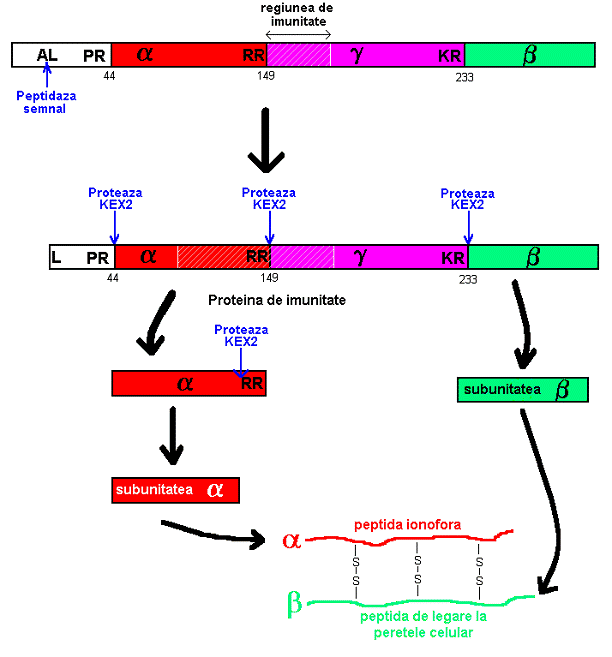 Fig. 1 Structura si maturarea preprotoxinei killer
Fig. 1 Structura si maturarea preprotoxinei killer
|
Politica de confidentialitate |
| Copyright ©
2025 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |