
DEZVOLTAREA SISTEMULUI NERVOS IN PRIMELE 72 ORE DE INCUBATIE
1. N EU RULATI A
In timpul gastrulatiei, interactiile dintre mezodermul dorsal si ectodermul supradiacent induc formarea tubului neural echivalent. Transformarea fiecarui strat de celule ectoderme in tub neural se numeste neurulalie si modificarile care au loc in embrion, neurula. In urma acesti proces, ectodermul initial este divizat in mai multe componente:
celulele ce vor migra in tubul neural si vor forma nevraxul;
celule aflate la nivelul epidermei pielii; celulele crestelor neurale care migreaza in regiunea de contact dintre tubul neural si epiderma.
La embrionul de pasari si mamifere, neurulatia are loc in doua faze:
neurulatia primara , in urma careia se formeaza encefalul si maduva spinarii pana la nivelul lombo sacral superior;
neurulatia secundara. in urma careia rezulta segmentele lombo - sacrale ale maduvei spinarii. Aceasta faza incepe in momentul in care neurulatia primara este aproape completa si implica agregarea celulelor mezenchimale din mugurele caudal intr-o coarda epiteliala longitudinala - coarda medulara - care se va cavita si va forma tubul neural caudal.
Neurulatia primara decurge in patru etape:
a. formarea placii neurale:
b. modelarea placii neurale;
c. formarea santului neural;
d. inchiderea santului neural:
1. 1. MECANISMUL NEURULATIEI
A. FORMAREA PLACII NEURALE
Placa neurala reprezinta primordiul sistemului nervos central. Ea este rezultatul inductiei epiblastului de catre mezodermul (eventual si endodermul) notocordal si nu devine distincta pana cand nu se formeaza procesul cefalic si mezodermul adiacent. Implica modificari ale formei celulelor ectodermale si expresia markerilor moleculari specifici. Nu se cunoaste inca stadiul exact in care apare (in stadiul de linie primara definita existand deja celule cu markeri caracteristici), localizarea pe harta epiblastica prospectiva si nici modul de transmitere a semnalului inductiv (vertical, orizontal).
Markerii moleculari identificati in cursul formarii placii neurale sunt:
N - CAM - in placa neurala, absent in ectodermul adiacent;
N - Cadherina - cu aceeasi localizare ca si N - CAM;
E - Cadherina - lipseste in placa neurala; prezenta in ectodermul adiacent.
Expresia N - CAM si N - Cadherinei in placa neurala se face sub influenta inductiva a mezodermului subdiacent.
B. MODELAREA PLACII NEURALE
Inaintea liniei primitive, viitoarele celule neurale ocupa probabil o arie semicirculara chiar in fata centrului blastodermului, unde se va forma nodul Hensen. Pe masura ce linia primitiva se formeaza, placa, neurala se alungeste devenind ovoida. in stadiul de linie primitiva definita, doar partile anterioare si laterale ale viitoarei placi neurale sunt asezate in afara blastodermului; material median, care va deveni podeaua tubului neural se dezvolta din marginea anterioara a nodului primitiv, de-a lungul procesului cefalic. cu care este asociat intim (Schoenwolf G.C.1981). in spatele nodului primitiv, portiunea centrala a placii neurale este nediferentiata si apare in fata liniei primitive ca o regresie a acestuia din urma. Oricum cu mult inainte ca linia primitiva sa dispara, cea mai mare parte a placii neurale, pe directie antero - posterioara .
Astfel, stadiile succesive pot fi studiate pe sectiuni ale aceluiasi embrion
sectiunea anterioara reprezinta tubul complet;
o sectiune mai posterioara reprezinta placa neurala pliata
iar o sectiune si mai posterioara reprezinta partea centrala a placii medulare ce dispare in masa nediferentiala a liniei primitive.
Modelarea placii neurale implica trei manifestari caracteristice:
cresterea diametrului apico - bazai la majoritatea celulelor neuro - epiteliale ce compun epiteliul columnar pseudostratificat al placii neurale.
- ingustarea transversala a placii neurale datorita scaderii diametrului transversal al celulelor (prin cresterea diametrului apico - bazai) si rearanjari celulelor neuroepiteliale prin fenomene de intercalare latero - mediala;
- Alungirea longitudinala a placii neurale datorita intercalarii latero - mediale a celulelor si diviziunilor celulare orientate.
Toate aceste modificari au ca substrat modificari ale formei, pozitiei si numarului celular neuroepiteliale
C. FORMAREA SANTULUI NEURAL
Acest proces include:
adancirea axiala a placii neurale;
formarea cutelor (crestelor) neurale.
CI. Adancirea placii neurale
Imediat dupa formarea pliului cefalic. partea centrala a placii neurale se scufunda sub forma unui sant santul neural. incepe imediat dupa capatul anterior al placii. Adancirea placii neurale este determinata de mecanisme intrinseci fiind rezultatul modificarii formei celulelor neuroepiteliale din columnara in piramidala (in pana de despicat), concomitent cu scaderea inaltimii lor. Ba a asociat cu formarea a trei regiuni speciale:
una mediana MHP (Median Hinge Point);
doua dorsalaterale DLMPs (Dorso Lateral Hinge Points ).
Cele doua regiuni ale neuroepiteliului si situate intre MHP si DLHPs de fiecare parte se numesc zone laterale L .
MHP se formeaza la nivelul mezencefalului si se intinde pe toata lungimea maduvei spinarii, reprezentand regiunea mediana, la nivelul careia placa neurala se ancoreaza de notocord si unde are loc adancirea placii neurale.
DLHPs apar la nivelul encefalului si a extremitatii caudale a tubului neural primar (in regiunea neuroporului posterior), fiind formate de neueroepiteliul dorsolateral si de rpiteliul nonneural adiacent. Fie reprezinta deci regiunile laterale ale placii neurale, unde aceasta se ancoreaza la ectodermul nonneural.
C.2. Formarea cutelor neurale
Marginile partii anterioare ale placii medulare se inalta oarecum deasupra blastodermului inconjurator . formand cute neurale. Formarea cutelor neurale este centrata in jurul celor trei zone balama. Ea implica mecanisme extrinseci, care determina a prima rotatie spre dorsal a cutelor neurale (cu punct fix in MHP) ceea ce duce la ridicarea lor. Urmeaza apoi o rotatie orientala medial, in jurul DLHPs care are ca rezultat convergenta cutelor neurale.
Cutele neurale cresc in marime si devin penate prin modificari la nivelul microtubulilor si m i cro fi 1 amentel or.
Colchicina. inhibatelor al polimerizarii microtubulilor inhiba elongarea acestor celule ca si citochalazina B (inhiba formarea microfilamentelor). Dupa inceperea adancimii, fiecare balama are rol (inhiba formarea microfilamentelor). Dupa inceperea adancimii, fiecare balama are rol de pivot orientand rotatia celulelor din jurul ei si placa neurala se indoaie. Ectodermul supradiacent e forta extrinseca e determina curbarea placii neurale spre interiorul embrionului. Lateral de placa neurala se formeaza trei tesuturi: epiteliul de suprafata (se continua medial cu placa neurala);
mezodermul (placa laterala):
endoderm ui (va forma intestinul).
indepartarea mezodermului si endodermului situate lateral de plac neurala nu impiedica formarea cutelor neurale, ceea ce demonstreaza ca numai epiteliul de la suprafata (ectodermul) si matricea sa extracelulara sunt implicate in acest proces, formarea cutelor neurale implica trei mecanisme extrinseci:
diviziunea celulelor ectoderme care determina cresterea volumului si suprafetei ectodermului;
intercalarea latero - mediala in plan transvers a celulelor; aplatizarea celulelor ectoderme.
Descrierea embrionului la 21-24 ore de incubatie - scanare la microscop electronic -
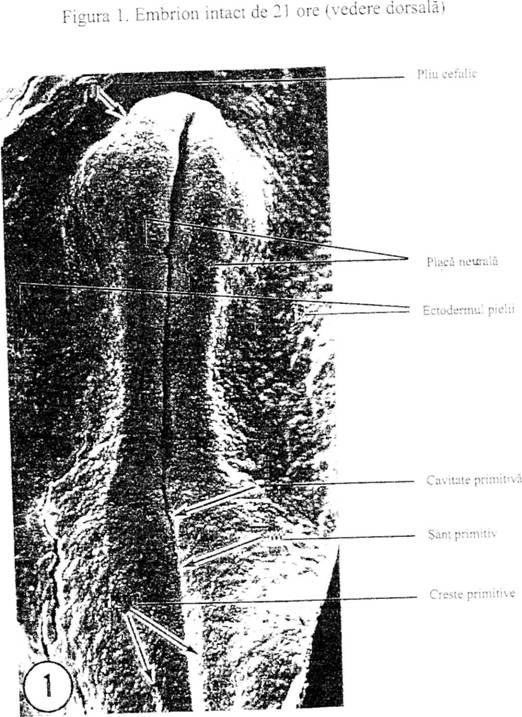
D. INCHIDEREA SANTULUI NEURAL
inchiderea santului neural este mediana de glicoconjugatele de pe suprafata celulelor, cere determina adeziunea celulara si fuziunea crestelor neurale. Adeziunea celulara se datoreaza sintezei de molecule de adeziune, iar fuziunea se realizeaza prin intercalarea prelungirilor citoplasmatice. Aceasta din urma incepe in regiunea mezencefalului in studiu de patru - cinci somite. in acest moment, marginile posterioare ale crestelor neurale nu au ajuns inca la nivelul primei somite. Dupa formarea unui prim punct de contact intre cele doua creste (cute) neurale, fuziunea acestora are loc in directia anterioara cat si posterioara.
Santul neural se inchide formand tubul neural cand crestele se unesc pe linie dorsomediana.
La capatul cefalic. peretele tubului e ingrosat si o serie de constrictii si dilatatii incep sa defineasca compartimentele cerebrale. in partea caudala a regiunii cefalice. tubul neural ramane ca un tub simplu ce se ingusteaza spre coada.
inchiderea completa a partii anterioare a tubului are loc in stadiul de 12 somite. Punctul final de inchidere a extremitatii anterioare a tubului neural se numeste neuropor anterior.
Neuroporul anterior este considerat de catre unii autori a reprezinta capatul anterior al axei neurale primitive, fiind astfel omolog cu neuroporul permanent de la Amphioxus. Aceasta ipoteza este inca discutabila avand in vedere modul de formare a extremitatii anterioare a encefalului. Acompaniind inchiderea tubului neural in aceasta regiune, pliurile neurale anterioare cresc rapid in continuare dincolo de capatul anterior al intestinului anterior. Planseul tubului neural nu ia parte insa Ia aceasta extindere, astfel incat varfurile crestelor neurale iau forma geniculara si se intind in fata capatului anterior initial al placii neurale. Capatul anterior al tubului neural prezinta la inchidere doua suturi: una dorsala si unde ventrala. Extremitatea suturii ventrale marcheaza marginea anterioara initiala a placii neurale si se intinde (in stadiul de zece somite) putin inaintea capatului anterior al placii orale.
De vreme ce sutura se intinde din apropierea infudibulului spre anterior, in jurul extremitatii frontale a encefalului si inapoi pana in regiunea pineala. exista o larga posibilitate de alegere a regiunii care sa fie numita neuropor. Exista un punct al suturii, situat langa capatul dorsal si este considerat ca fiind neuroporul anterior. Neuroporul astfel definit si intinde in fata capatului anterior actual al placii neurale primitive si este lipsit de importanta evolutiva. Sutura este locul de formare a laminei terminalis.
in partea posterioara. crestele neurale fuzioneaza mult mai rapid. Nivelul la care are loc unirea celor doua creste corespunde cu cel al formarii noilor somite. in partea posterioara a ultimei somite formate crestele neurale se indeparteaza una de alta, inaltimea lor scade si ele se unesc cu ectodermul lateral de linia primitiva. Deschiderea rezultata astfel de forma ovoida se numeste sinus romboidal. Aceasta zona se reduce pe masura ce se formeaza alte somite si in timp ce linia primitiva regreseaza , astfel incat in stadiul de 13 somite, sinusul romboidal apare sub forma unei despicaturi ovoide alungite cere se continua posterior cu santul liniei primitive.
Mai tarziu in stadiul de 20 - 21 somite, tubul neural complet inchis se uneste posterior cu nodul Hensen. Deschiderea care ramane inaintea inchideri complete a tubului neural este uneori numita neuroporul posterior, desi acest termen nu e adecvat. Adevarul neuropor posterior este reprezentat in momentul unirii crestelor neurale, de adancitura nodului Hensen [4].
1. 2. MISCARILE CELULARE IN EP1BLAST IN TIMPUL NEURULAT1EI
Celulele MHP
Celulele MHP precursoare jn timpul modelarii si cudarii placii neurale au patru tipuri de manifestari morfogenetice: diviziuni celulare;
modificari ale formei si marimii celulare;
modificarea pozitiei celulelor;
interactiuni intercelulare inductive,
Majoritatea celulelor MHP (70 %) sunt piramidale (apar in forma de pana de despicat), spre deosebire de celulele placii neurale initiale care sunt fusiforme in cea mai mare parte. Comparativ cu inaltimea celulelor placii neurale initiale care sunt fusiforme in cea mai mare porte. Comparativ cu inaltimea celulelor placii neurale initiale, inaltimea celulelor MHP este cu 25 % mai mica. Aceste modificari prezentate mai sus , sunt rezultatul interactiunilor inductive cu notocordul . In timpul neurulatiei apar intercalari latero - mediale a celulelor MHP care determina extensia convergenta a placii neurale.
Ciclul celular al celulelor MHP este de 12 ore cu 65 % mai lung decat al celulelor placii neurale inaintea formarii regiunilor MHP si L.

Celulele L
in timpul modelarii eudarii placii neurale, manifestarile morfogenetice ale celulelor L
sunt:
diviziunea celulara;
modificarea formei si marimii celulelor;
modificari ale pozitiei celulelor.
O mare parte a celulelor L (70 % ) sunt fusiforme la fel ca cele ale placii neurale initiale.
Celulele L au insa diametrul apico - bazai cu 30 % mai mare ca al celulelor placii neurale primitive. Celulele L isi modifica pozitia in timpul neurulatiei prin intercalarea latero -mediana , determinand extensia convergenta a placii neurale.
Ciclul celular al celulelor L este identic cu cel al placilor neurale inaintea formarii regiunilor MHP si L si are o durata de opt ore.
Celulele SE (Surface Epithelial)
a. Celulele SE cxtraembrionare
Celulele SE au cel putin doua tipuri de manifestari morfogenetice in timpul modelarii si eudarii placii neurale: diviziune celulara:
modificari in forma si marimea celulelor;
Ciclul celular este de zece ore, deci mai scurt ca cel al celulelor MHP si mai lung decat al celulelor L. In ceea ce priveste forma, aceasta se modifica din cuboidala in sevamoasa iar inaltimea lor scade cu circa 60%.
b. Celulele SE intraembrionara
Manifestarile lor morfogenetice din cursul neurulatiei sunt: diviziune celulara;
intercalare latero - mediana ce determina o miscare de convergenta - extensie; mentinerea formei si marimii initiale.
Formarea celulelor neurale e determinate de mecanisme intriseci ce realizeaza adancirea placii neurale si de mecanisme extrinseci (epiteliul de suprafata si matricea asociala ce determina ridicarea si convergenta cutelor neurale).
In cursul modelarii placii neurale rearanjarile celulare faciliteaza curbarea placii neurale, prin scaderea largimii si cresterea lungimii acesteia, permitand fortelor extrinseci sa ridice cutele neurale spre linia mediana unde vor fuziona [2].
2. CRESTELE NEURALE
Celulele crestale deriva din neuroectoderm si dau nastere mai multor tipuri celulare: neuroni si nevrogliile senzorilor, sistemului nervos simpatic si pareasimpatic; celulele producatoare de adrenalina din medulosupfarenala: componentele tesutului osos si conjunctiv al capului.
CAILE DE MIGRATIE A CELULELOR CRESTALE (GANGLIONARE) ALE TRUNCHIULUI
Concomitent cu definitivarea SNC se formeaza crestele ganglionare in unchiul neuroectoblastic, mai tarziu in segmentul cerebral al SN decat in cel medular. Sunt alcatuite din celule mici neuroectoblastice care migreaza spre marginile santului neural. Cand aceste regiuni se unesc, materialul celular crestai se contopeste dorsal fata de tubul neural. Materialul se desparte mai apoi in doua mase celulare matemerice.
a. Calea neurala
Migratia celulelor crestale are loc in acest caz prin portiunea anterioara a somotei, dand nastere neuronilor senzoriali simpatici, celulelor Schwann si celulelor medulosuprarenalei. O parte din aceste celule ajung la nivelul aortei dorsale unde formeaza ganglionii simpatici. Celulele crestale ce migreaza prin regiunea posterioara a somitei de-a lungul tubului neural anterior si posterior, vor patrunde apoi prin portiunea anterioara a somitei respective sau a celulelor adiacente. Ele se vor uni cu celulele situate initial in dreptul regiunii anterioare a somotei. formand aceleasi structuri.
b. Calea dorsolaterala
Migratia celulelor crestale pe aceasta cale are loc sub ectodermul embrionar si ele vor forma celulele pigmentare (melanocite).
CAILE DE MIGRATIE A CELULELOR CRESTALE CRANIALE
Celulele crestale craniale migreaza inainte de inchiderea tubului neural si dau nastere mezenhimului facial.
Celulele crestale originare din prozencefal si mezencefal contribuie la formarea procesului nazal, palatului si mezenhimului primului arc faringian.
Celulele crestale din regiunea anterioara a rombencefalului formeaza mezenhimului celui de-al doilea arc faringian.
Celulele crestale cervicale contribuie la mezenhimul arcurHor III. IV si VI.
DIFERENTIEREA TUBULUI NEURAL
De-a lungul tubului neural apar neuroepiteliul germinai unistratificat ale carui celule au muchiile situate la diferite nivele. Ca rezultat al proliferarii acestor celule se formeaza un strat secundar, mantaua (zona paleala) care se ingroasa, celulele diferentiindu-se la momente diferite in neuroblaste si gliohlaste. Neuroepiteliul germinai devine zona ventriculara si mai tarziu ependimara.
Tubul neural reprezinta deci primordiul encefalului si al maduvei spinarii. Cavitatea sa se transforma in ventriculii din encefal si in canalul ependimar al maduvei spinarii. in primele stadii de dezvoltare insa, neexistand o limita clara intre encefal si maduva. Aceasta apare odata cu formarea somitelor mezodermale pentru ca primele patru somite complet formate (neluand in seama somitele rudimentare) sunt cefalice. Prin urmare tot tubul neural din fata celui de-al cincilea sant intersomatic este cranial.
1. FORMAREA REGIUNII CEREBRALE
La nivel anatomic lumenul tubului neural se mareste datorita acumularii de lichid si nu pe baza ingrosari tesuturilor.

Descrierea embrionului la 24 ore incubatie
Fisura 1. Vedere in ansamblu

Neuropor cranial Proamnios Tub neural
Extinderea caudala a pungii subcefalice
|
,inie primitiva |
Cute neurale Somite
Dupa 30 de ore de incubatie, datorita inchideri neuroporului cranial si a presiunii lichidului asupra peretilor tubulari se delimiteaza net vezicula cerebrala. Tesuturile dorsale din jur produc constructia tubului separand regiunea cerebrala prezumtiva de viitoarea coarda spinala. Dilatarea veziculelor are loc lent iar regiunea ocluzata a tubului se redeschide dupa largirea initiala rapida a ventriculelor cerebrale.
inainte de formarea maduvei spinale, vezicula cerebrala se diferentiaza in trei segmente: preozencefalul - prezinta rudimentele veziculare optice si o sutura in partea anterioara a planseului datorita modului sau de formare;
mezencefalul - ce apare separat de prozencefal datorita unei constructii ce apare in stadiul de 6 - 7 somite;
rombencefalul - ce se prezinta separat de mezencefalul printr-o constrictie ce apare in stadiul de 9 - 10 somite. iar jumatatea sa anterioara se caracterizeaza in stadiul de 12 sonite prin existenta a cinci diviziuni (neuromere sau rombomere) separate prin constrictii.
Rombomerele sunt izolate de cele vecine. La embrionul de gaina, primii neuroni apar in rombomerele n. n, r(>.
Neuronii gliali din: - n formeaza nervul V cranial;
- n formeaza nervii VI si VIII; -re formeaza nervul IX. Dupa 48 ore de incubatie prozencefalul se diferentiaza in telencefal (anterior) si diencefal, rombcncefal in metencefal si mielencefal, iar mezencefalul ramane nediferentiat. Punctele de referinta morfogenetice in intervalul de 37 - 72 ore de incubatie sunt: lamina terrrminalis - peretele anterior al tubului neural;
infundibil - situat in partea anterioara a notocordului;
spatiul intersomotic 5 - care marcheaza partea posterioara a encefalului [2].
EVOLUTIA TELENCEFALULUI
Telencefalul in generarea emisferelor cerebrale se structureaza in doua directii diferite: vertical (pe strate) si orizontal (pe regiuni).
Anumiti nuroblastici din zona paleala vor genera o zona secundara neuronala numita cortex neuropaleal, deasupra substantei alese. Aceasta e format din sase strate care contin
Figura 2. Ansamblu
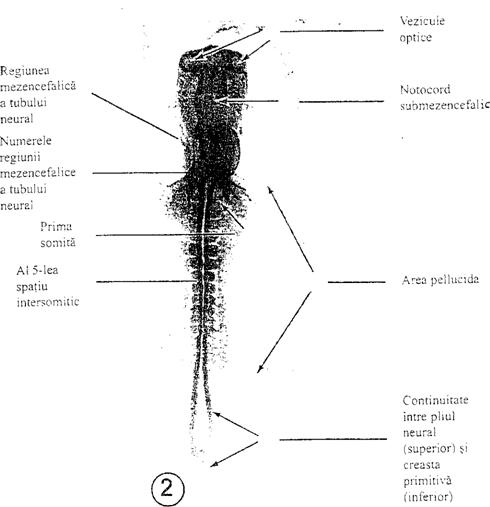
corpii neuronalii. Stratele difera intre ele prin proprietati functionale, tipul neuronilor si conexiunile realizate.
De asemenea, organizarea orizontala a cortexului cerebral implica peste 40 regiuni corespunzatoare unor diferite procese.
Dupa ultima mitoza, cei mai noi neuroni formati, migreaza radial de-a lungul procesului glial din zona ependimara formand sub pia mater placa corticala. O singura celula mama a stratului ventricular poate produce toti neuronii si gliile pentru un strat cortical.
Stabilirea identitatii laminare (stratul in care migreaza fiecare neuron) se face abia in timpul diviziuni finale. Astfel celulele din stratul sase al unui encefal tanar, implantate in stratul doi al unui encefal adult, dupa ultima diviziune migreaza numai in stratul sase. In cazul in care insa sunt implantate inainte de diviziunea finala, vor migra in stratul doi.
Prin metoda colorarii s-a demonstrat ca 80 % din neuronii tineri migreaza radial pe procesul glial in placa corticala si 12 % migreaza lateral dintr-o regiune coiticala in alta. Specializarea functionala a regiunilor corticale apare dupa neurogeneza.
EVOLUTIA DIENCEFALULUI
Diencefalul se formeaza prin proliferarea neuroblastelor si glioblastelor foarte intensa in peretii laterali astfel incat de o parte si de alta a ventricolului III se organizeaza doua mase celulare: talamusul si corpii geniculati laterali.
Peretele neural al diencefalului va forma veziculele optice primitive, chiasma optica si regiunea infundibulo - tuberiana (tuber).
Primordiile optice mut doua formatiuni sferice atasate de prozencefal printr-un pedicul scurt (pedicul optic) inainte de aparitia diencefalului, iar dupa aparitia acestuia raman prinse definitiv de portiunea anterolaterala a bazei diencefalului.
Lobul nervos al hipofizei (retrohipofiza) si corpii mamilari se dezvolta din tuber sub forma unor invaginatii diverticulare.
Glanda pineala sau epifiza si membrana tectoria a ventricolului III se formeaza din peretele dorsal si se ingusteaza si se ingusteaza formand cele doua orificii Motiro.
EVOLUTIA MEZENCEFALULUI
Mezencefalul este portiunea encefalului care ocupa varful curburii craniului, curbura formata in mare parte datorita cresteri rapide a plafonului mezencefalului.
El prezinta un planseu scurt si un tavan lung si arcuit iar in primele 72 ore de incubatie peretii sai au o grosime uniforma. Dupa stadiul de 36 somate, peretii laterali se evagineaza rapid formand lobii optici.
EVOLUTIA ROMBENCEFALULUI
Rombencefalul este portiunea posterioara a creierului primitiv si va da nastere metencefalului si mielencefalului. Metencefalul va forma cerebelul si puntea iar mielencefalul va forma bulbul rahidian. Metencefalul este scurt, include doar primele ncuromere rombencefalice si pana la 72 de ore tavanul sau are aceeasi grosime cu al mezencefalului. Tavanul mielencefalului este subtire iar peretii laterocentrali si planseul sunt ingrosate. Neuromerele raman distincte dar isi modifica forma.
EVOLUTIA CEREBELULUI
Anumiti neuroni din substanta alba se diferentiaza in nudei, relee intre stratele extreme cerebeloase si alte zone cerebrale. Alta zona a neuroblastilor (neuroni Purkinje diferentiati) va migra la suprafata externa formand stratul germinai extern care prolifereaza intens la suprafata. Compartimentele acestui strat contin celule granulare. precursorii majoritatii neuronilor cortexului cerebelos. Celulele granulare migreaza inapoi in substanta alba formand stratul granular intern. Neuronii Purkinje sunt generati de stratul ependimar. Unul singur poate forma 100 000 sinapse.
2. FORMAREA MEDUVEI SPINARII
Peretii medulari primitivi sunt alcatuiti din celule prismatice dispuse intr-un singur strat. Peretii ventrali si dorsali raman ca lame subtiri bazale si respectiv dorsala, iar peretii laterali incep sa se ingroase. Placa fundamentala se afla anterolateral iar placa alara posterolaterala.
In jurul lumenului epiteliul primitiv prolifereaza rapid formand zona germinativa cu elemente celulare care vor diferentia in neuroblaste si glioblaste ce vor constitui zona paleala (mantaua). La suprafata se constituie valul marginal sarac in elemente celulare care insa se dezvolta foarte mult pana cand la acest nivel se vor forma cordoanele medulare.
Substanta cenusie se dezvolta mai mult in coloana anterioara decat in coloana posterioara pentru ca activitatea proliferativa a fost mai intensiva in dreptul placii fundamentale decat in dreptul alarei.
Santul limitat intern se formeaza pe fata interna a peretilor laterali si divide maduva in jumatatea dorsala si ventrala. Materialul medular situat ventral fata de sant va forma neuronii somato- si visceromotorii din cordoanele anterioare medulare, iar cel situat dorsal fata de sant se organizeaza in neuronii somato- si viscerosenzitivi ai coloanelor posterioare.
Elementele mezenchimale din jurul maduvei vor organiza meningele: pia mater, arahnoida si dura mater.
DEZVOLTAREA ORGANELOR DL SIMT
DEZVOLTAREA VEZICULELOR OPTICE
In stadiul de sant ventral, lateral, se dezvolta gropitele optice ca doua excrescente care dupa constituirea celor trei veziculare cerebrale devin vezicule optice care printr-un pedicul scurt sunt anexate peretilor laterali ai prozcncefalului. Dupa aparitia diencefalului, veziculele raman anexate fetelor lateroventrale la granita diencefalului cu telencefalul.
Veziculele cresc in dimensiuni datorita proliferarii celulare iar peretii lor se ingroasa ajungand in vecinatatea ectoblastului adiacent. Prin inductie celulara ectoblastice devin prismatice generand placodele lenticulare care se transforma rapid in gropite lenticulare ale caror margini se apropie concentric si formeaza veziculele lenticulare. Dupa separarea de ectoblastul de origine, incepe adancirea lor in peretii veziculelor optice, acestea devenind cupe optice cu pereti dubli si induc formarea corneei. Apare si o ingrosare inegala a peretilor:
Peretele intern se pl uri stratifica organizand retina primitiva iar peretele exterior ramane un epiteliu cubic monostratificat si va genera stratulpigmentar retinian. Spatiul dintre ei dispare treptat.
Corpii neuronali pigmentari se evagineaza si sintetizeaza organite specializate caracterizate ca forma si marime fiecarei celule. Membrana evaginatiilor se pliaza in forma de sac in care se vor deplasa fotoreceptorii.
Curbarea corneei e controlata de presiunea lichidului intraocular.
Sub influenta retinei neurale celulele interne ale veziculei lenticulare se alungesc si produc fibrele cristaliniene. Celulele anterioare lenticulare constituie epiteliul germinai al carui diferentiere in fibre lenticulare dureaza foarte mult.
Dintr-o regiune a cupei optice se formeaza irisul, iar invelisurile globilor oculari se formeaza din mezenchimul condensat in jurul veziculelor optice organizand coroida. sclerotica si tesutul lamelar al corneei. Epiteliul anterior al corneei este insa de natura ectoblastica.
DEZVOLTAREA URECHII INTERNE
Elementele ectoblastice devin prismatice in vecinatatea peretilor laterali ai romencefalului si se pluristratizeaza formand placodul auditiv al carui centru se invagineaza si genereaza fosetet acustica. Prin inchiderea concentrica a fosetei se formeaza vezicula optica primitiva (otocistul).
DEZVOLTAREA ORGANULUI OLFACTIV
Placodele olfactive apar ca doua ingrosari ectodermice de o parte si de alta a stomodeumului. De fiecare parte a acestora mezodermul ridica ectodermul inducand invaginerea placodelor si formarea gropitelor olfactive si mugurele nazal extern. In urma acestui proces se formeaza fósetele olfactive adancite in santurile delimitate ale mugurilor
nazali externi si a fisurilor nazale, in ansamblu formand campurile nazale Hiss. P rin diferentierea celulelor placodului olfactiv se formeaza pituitara, organul olfactiv propriu-zis, iar din materialul primitiv al acestuia apar celulele epiteliale de sustinere si celulele Schultze, viitoarele celule mitrale.
|
Politica de confidentialitate |
| Copyright ©
2025 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Conceptul de mimesis |
| DEZVOLTAREA MEZODERMULUI IN PRIMELE 72 DE ORE DE INCUBATIE |
| Metodele si tehnicile de studiu a documentelor |
| DEZVOLTAREA SISTEMULUI NERVOS IN PRIMELE 72 ORE DE INCUBATIE |
| Termeni si conditii |
| Contact |
| Creeaza si tu |