Fiziologia celulara sau citofiziologia studiaza fenomenele vietii ce se petrec in interiorul celulei. Ea constituie capitolul fundamental al fiziologiei, deoarece toate manifestarile vitale au loc in materia vie celulara. Din acest motiv cunoasterea vietii celulare poate oferi cheia tuturor manifestarilor fiziologice de care sunt capabile organismele vii.
Celula poseda toate insusirile unui organism viu, capabil sa duca o viata independenta. Acest lucru este dovedit nu numai prin existenta organismelor monocelulare, ci si prin posibilitatea de a recolta celule din organismele pluricelulare care, cultivate pe medii artificiale, cresc, se inmultesc si dau nastere la plante intregi; s-au obtinut astfel experimental culturi de celule, pornindu-se de la o singura celula sau de la un fragment minuscul de tesut.
Celula este un sistem termodinamic deschis; ea face schimb permanent de substante cu mediul in care traieste, transforma substantele asimilate si energia in procesul de metabolism.
Forma celulelor vegetale este foarte diferita: forma sferica se intalneste la bacterii (coci); forma poliedrica-izodiametrica, apropiata de cea sferica este caracteristica tesutului meristematic din varfurile de crestere ale radacinii si tulpinii; celulele din tesuturile liberiene sunt fuziforme, iar cele din tesuturile conducatoare, mecanice si in unele tesuturi absorbante si secretoare (laticiferele) sunt tubulare.
Dimensiunile celulelor vegetale variaza intre 10 μm si 50 cm (fibrele liberiene) sau chiar mai multi metri (laticiferele nearticulate de la unele specii din familia Euphorbiaceae), valorile medii fiind de aproximativ 10 - 100 μm.
Morfologia externa a celulelor vegetale este rezultatul influentei conditiilor de viata de care beneficiaza celulele in cadrul tesutului din care fac parte. In acelasi timp, forma si marimea celulelor corespund functiei fiziologice pe care acestea trebuie sa o indeplineasca in tesutul respectiv.
Celula procariota - a aparut - ca organism elementar - cu 3,5-4 miliarde de ani in urma. Celulele procariote nu contin organite bine diferentiate de tipul nucleului, mitocondriilor, cloroplastelor etc. Informatia genetica este inclusa in unicul cromozom, care consta dintr-un lant de macromolecule de ADN, sub forma de inel. Continutul celulei consta din citoplasma densa si o regiune nucleara traversata de macromolecula de ADN.
La cianobacterii intalnim membrane sub forma de inele, ce participa la procesul de fotosinteza. Aceste membrane contin in stare difuza clorofila 'a' si ficocianina (pigmentii implicati in procesul de fotosinteza).
Celula procariota mai contine ribozomi - organite raspunzatoare de sinteza proteinelor. Nu s-au evidentiat alte structuri interne, ceea ce nu inseamna insa ca aceste celule sunt lipsite de posibilitati biochimice multiple. Din contra, s-au observat la acest nivel 500-1000 reactii biochimice.
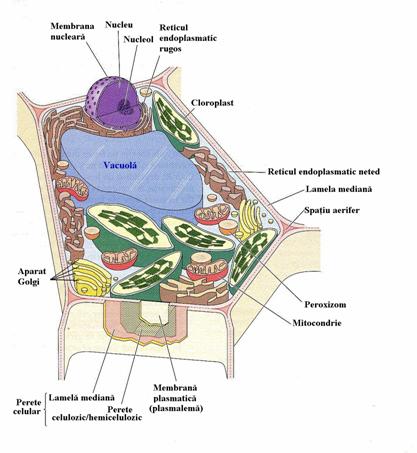
Celula eucariota este de cateva ori mai mare, comparativ cu celula procariota, fapt ce constituie o dovada in plus ca ea reprezinta o etapa superioara in evolutia generala a sistemelor vii (vezi fig. 3 si 4; fig. 8).
Marirea dimensiunilor celulelor, precum si perfectionarea structurii lor au fost insotite de cresterea cantitatii de ADN, care a dus la cresterea si perfectionarea unei arhitecturi complet noi de distribuire a materialului ereditar.
Pentru celula eucariota este caracteristica o organizare mult mai complexa decat pentru cea procariota, din care probabil a provenit. In ea intalnim deja 'o distribuire a functiilor', datorita diferentierii macro- si microstructurilor celulare.
Celula este acoperita cu o membrana citoplasmatica (plasmalema), iar celula vegetala mai dispune inca de o anvelopa celulara (de natura celulozica), numita perete.
Partea principala a celulei o constituie continutul intern - protoplasma - in care au loc cele mai importante procese vitale.
Protoplasma unei celule dintr-un tesut viu comunica cu protoplasma celulelor invecinate, a caror totalitate formeaza organismul pluricelular. Comunicarea se realizeaza prin plasmodesme (fire protoplasmatice).
Protoplasma consta din citoplasma, in care se afla organite celulare: nucleul, plastidele, mitocondriile etc., organite ce pot fi observate la microscopul optic. Alaturi de acestea, in hialoplasma mai exista o serie de structuri submicroscopice cum ar fi: aparatul Golgi, reticulul endoplasmatic, ribozomii, microtubulii etc., fiecare organit celular avand o structura si o ultrastructura specifica si, bine inteles, functii specifice.
Caracteristice pentru celula vegetala matura sunt, de asemeni, vacuolele sau vacuola unica, pline sau plina cu suc celular, la care se adauga diferite incluziuni solide (granule de aleurona, granule de amidon s.a.).
Functiile organitelor celulare se studiaza 'in vitro', dupa separarea lor prin metoda ultracentrifugarii.
Citoplasma (vezi fig. 3, 4, 8) este masa fundamentala a protoplasmei vii, in care sunt incluse organitele celulare. Ea este componenta esentiala a celulelor vii, aflandu-se in stare fluida si in neintrerupta miscare; coagularea sa determina incetarea manifestarilor vitale si, implicit, moartea celulei.
Substantele organice ce constituie elemente de constitutie ale citoplasmei sunt: proteine hidrofile, fosfolipide, acizi nucleici etc.
Marea varietate de proteine existente in citoplasma determina functii metabolice si enzimatice caracteristice, ce imprima specificitate.
In celulele tinere spatiul citoplasmatic este cuprins intre membrana plasmatica externa (plasmalema) si invelisul nuclear.
|
|
In celulele mature, cu vacuole mari, citoplasma reprezinta un strat subtire, cuprins intre membrana plasmatica externa (tonoplastul), ce se margineste cu anvelopa celulara celulozica (peretele) si cea interna, ce se margineste cu vacuola. Prima membrana plasmatica, care regleaza patrunderea si iesirea substantelor din celula a fost denumita de catre W. PFEFFER, in 1877, plasmalema, iar membrana plasmatica ce inconjura vacuolele celulelor vegetale a primit denumirea de tonoplast.
|
Fig. 8- Celula din mezofilul foliar prezentand principalele sisteme membranare si peretele celular (d. Buchanan si colab., 2000) |
Alaturi de membranele plasmatice amintite, in componenta citoplasmei intra hialoplasma cu organitele citoplasmatice.
Hialoplasma este o masa transparenta, omogena si aparent lipsita de structura, numita si matrice citoplasmatica.
Plasmalema (vezi fig. 3 si 8) are proprietati selective. Ea controleaza activitatea celulei, absorbtia si excretia unor substante si joaca un rol important in procesul de nutritie a celulei cu ioni minerali, prin schimb ionic.
Tonoplastul (vezi fig. 3 si 8) inconjura vacuolele celulelor vegetale si este o membrana plasmatica de tip biologic, lipoproteica, ca si plasmalema. El se deosebeste insa de plasmalema prin structura, compozitie chimica cantitativa si prin gradul diferit de permeabilitate.
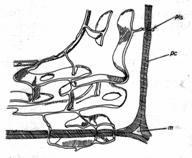
Reticulul endoplasmatic (R.E.) (fig. 9) este un sistem complex de cavitati (sacule, cisterne, vezicule sau tubuli ori canalicule fin ramificate si anastomozate., iar in celulele mai varstnice un sistem de 'cisterne' turtite si latite) in hialoplasma, toate delimitate de citosol prin membrane simple, elementare, lipo-proteice, asemanatoare ca structura cu plasmalema.
|
|
|
Fig. 9 - Schema ultrastructurii reticulului endoplasmatic: m. - meat; p.c. - perete celular; pls. - plasmodesme (d. Buvat, 1969) |
Lumenul cisternelor si tubulilor comunica cu spatiul perinuclear al anvelopei nucleare si cu porii acesteia.
In membranele R.E. neted (vezi fig. 3 si 8) se formeaza hidrati de carbon, lipide, terpenoizi, iar in cel granular (vezi fig. 3 si fig. 8) proteine, enzime, substante absolut necesare procesului de sinteza a produsilor de tipul polizaharidelor, din componenta peretelui celular etc.
|
|
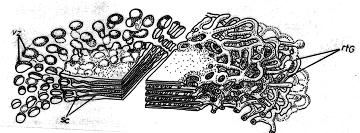
Aparatul Golgi (fig. 10), numit astfel in memoria omului de stiinta italian care l-a descoperit, este alcatuit din unitati morfologice denumite dictiozomi. Acestia sunt formatiuni lamelare, formate din 2-10 sacule sau cisterne, cu aspect de discuri suprapuse, aproximativ circulare, unite la centru.
|
Fig. 10 - Schema ultrastructurii unui dictiozom: r.t.G. - reteaua trans Golgi; sc. - sacule; vz. - vezicule (d. Kaussmann, 1989) |
Din punct de vedere functional, aparatul Golgi este legat de activitatea secretoare a protoplasmei.
Ribozomii (vezi fig. 3 si 8) sunt particule ultrastructurale de natura ribonucleoproteica, lipsiti de membrana, cu dimensiuni de 150-300 Å; au fost evidentiati de savantul roman G. E. PALADE in 1953 in omogenatele hepatice si in ultrasectiuni celulare si de catre E. ROBINSON si R. BROWN in celulele radacinii de bob. Ribozomii sunt raspanditi in toata celula (in citoplasma): la suprafata nucleului, in interiorul cloroplastelor si al mitocondriilor sau chiar se afla liberi in citoplasma.
Functia fiziologica principala a ribozomilor este legata de participarea lor in procesul de sinteza a proteinelor.
Ribozomii liberi participa la sinteza proteinelor implicate in procesul de diferentiere celulara, la sinteza unor proteine cu functii specifice, precum si la sinteza enzimelor proteolitice.
Ribozomii aderenti la membranele R. E. de tip rugos sau pe fata externa a membranei nucleare sunt implicati in procesele de sinteza a proteinelor destinate secretiei celulare.
|
|
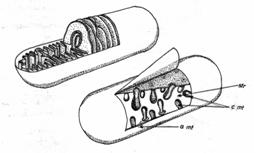
Condriozomii (fig. 11) - sunt cunoscuti azi ca principalii centri energetici (de respiratie) ai celulei.
In general, condriozomii sunt pre-zenti atat in celulele vegetale, cat si in cele animale dar lipsesc la bacterii.
|
Fig. 11 - Schema ultrastructurii mitocondriei: a.mt. - anvelopa mitocondriei; c.mt. - creste mitocondriale; str. - stroma mitocondriala (d. Kaussmann., 1989 ) |
In condriozomi se acumuleaza cele mai mari cantitati de A.T.P. din celula si tot aici are loc fenomenul de oxidare a metabolitilor, proces ce furnizeaza energia necesara transformarilor metabolice.
Numarul condriozomilor dintr-o celula este proportional cu intensitatea metabolismului acesteia.
Condriozomii reprezinta deci 'uzina biochimica (energetica)' a celulei, in care se elibereaza energia incorporata in substantele energetice de natura organica (in etapa finala a ciclului Krebs), energie ce se acumuleaza in legaturi macroergice fosfatice de tip ATP, prin procesul de fosforilare oxidativa. ATP-ul reprezinta sursa universala de energie a celulei.
Lizozomii (vezi fig. 3 si 8) sunt organite citoplasmatice sferice sau ovoidale, cu o membrana lipo-proteica simpla, asemanatoare cu plasmalema. Ei sunt bogati in enzime, in special hidrolaze acide. Intervin in hidroliza substantelor de rezerva din celulele endispermului, in procesul de germinare al semintelor.
Peroxizomii se intalnesc in numar mare in celulele mezofilului foliar, fiind dispusi frecvent alaturi de cloroplaste, cu care se pare ca vin chiar in contact.
Functia peroxizomilor consta in aceea ca peroxidaza si catalaza continute in ei scindeaza H2O2 in O si H2O si prin aceasta organitele respective intervin in procesul de respiratie celulara, atat in celulele mezofilului plantelor, cat si in celulele ficatului si rinichiului la mamifere.
Glioxizomii isi manifesta activitatea in momentul germinarii semintelor bogate in grasimi, cand apar in aceste seminte numerosi glioxizomi ce contin enzime specifice ciclului acidului glioxilic (izocitrat liaza si malat sintetaza); ele participa in mod activ la procesul de transformare a grasimilor in glucide, intr-un proces de glicoliza inversa (gluconeogeneza). De asemenea, ei reprezinta principalul organit celular in care are loc biodegradarea acizilor grasi, prin procesul de β - oxidare.
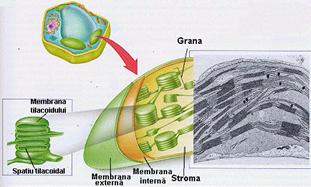
Plastidele sunt organite permanente de mare importanta pentru celula vegetala (vezi fig. 3 si 8). Totalitatea plastidelor incluse in citoplasma celulei formeaza plastidomul celular.
Plastidele lipsesc din celulele animale si din unele cianobacterii si bacterii, dar chiar si din cele ale unor macroorganisme (ciuperci si chiar plante superioare).
In anul 1883 A. SCHIMPER a descris cele trei tipuri de plastide existente in celula, tinand cont in special de absenta sau prezenta culorii: leucoplastele - plastide incolore, cloroplastele - plastide verzi si cromoplastele (sau carotenoplastele) - plastide colorate (in alte culori decat verde).
a) Cloroplastele sunt organite specializate, cu rol in procesul de fotosinteza, raspandite in toata masa citoplasmei, prezente doar in celulele vegetale.
La microscopul optic apar ca niste grauncioare verzi ovoidale, turtite; uneori au aspect de haltera.
Marimea lor este relativ constanta pentru acelasi tip celular, aparand insa deosebiri sexuale, genetice sau ecologice, in legatura cu acest parametru. Asa de exemplu ele sunt mult mai mari la plantele de umbra, dar si continutul lor in clorofila este mai mare, comparativ cu indivizii crescuti in lumina intensa.
Ultrastructural, cloroplastul este format din (fig. 12):
anvelopa plastidiala - formata din doua membrane elementare, separate de un spatiu clar;
stroma sau matricea - ce constituie substanta fundamentala cu ribozomi, ADN, granule de amidon, picaturi lipidice in ea;
sistemul de lamele
incluziuni reprezentate prin globule lipidice si granule de amidon.
|
|
|
Fig. 12 - Schema ultrastructurii cloroplastului (d. Berg, 1997) |
Cloroplastele reprezinta sediul sintezei primare a substantelor organice in procesul de fotosinteza si au o anumita autonomie in interiorul celulei, datorita ADN-ului propriu.
Conform parerii unor cercetatori, cloroplastele au fost bacterii fotosintetizante care, devenind simbionte intracelulare, au conferit celulelor capacitatea de fotosinteza ('Teoria Seriala a Endosimbiozei').
b) Cromoplastele au o structura asemanatoare cu cea a cloroplastelor si sunt colorate in galben, portocaliu, rosu sau in nuante intermediare, datorita prezentei in stroma lor a unor pigmenti din grupa carotenoizilor: carotenul - portocaliu, xantofila - galbena; licopenul - rosu etc. Din combinarea acestor pigmenti rezulta culorile specifice ale petalelor florilor, ale fructelor, frunzelor sau ale unor radacini.
c) Leucoplastele sunt plastide incolore, intalnite in celulele meristematice, in citoplasma sporilor si a gametilor femeli, in seminte, tuberculi, radacini si in majoritatea organelor incolore la angiosperme.
Leucoplastele se intalnesc, de asemeni, la plantele superioare adaptate la parazitism, care au pierdut proprietatea de fotosinteza, precum si in algele diatomee marine.
Sub influenta luminii ele sunt capabile sa formeze structuri tilacoidice si sa capete culoarea verde.
Dintre alte plastide, amintim: amiloplastele (in care se depoziteaza ca substante de rezerva amidonul), oleoplastele (in care se depoziteaza substante grase ) sau proteoplastele ( in care se depoziteaza proteine).
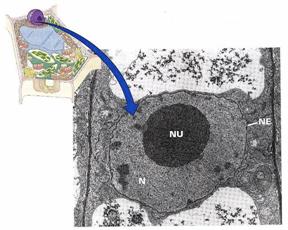
Nucleul (vezi fig. 3 si 8; fig. 13) descoperit in 1833 de catre BROWN, se considera astazi ca este un component prezent permanent in toate celulele vegetale si animale.
|
|
Celulele care au nucleu difuz, neindividualizat strict, se numesc procariote, iar celulele cu nucleu bine individualizat se numesc eucariote.
Rolul fundamental al nucleului in viata celulei este de a transmite caracterele ereditare in timpul diviziunii celulare si de a coordona functiile vitale ale acesteia..
|
Fig. 13 - Imagine electron microscopica a nucleului celular (N) la celule de Vicia faba L., evidentiind membrana nucleara dubla (NE) si nucleolul situat central (NU) (d. Buchanan si colab., 2000) |
Vacuola (vezi fig. 3, 6, 8) este o componenta nevie a celulei, plina cu suc vacuolar, deosebit de important pentru celula vegetala adulta.
Celulele meristematice sunt lipsite de o vacuola unica si voluminoasa, vizibila la microscopul fotonic; aici ele apar foarte mici, vizibile doar la microscopul electronic, dupa o colorare cu coloranti vitali.
In procesul cresterii si dezvoltarii celulei vegetale se observa formarea de vacuole mari; pe masura maturizarii celulelor, vacuolele mici se contopesc, se maresc in volum, iar la sfarsitul procesului de diferentiere se formeaza o vacuola mare centrala, care ocupa aproape toata cavitatea celulara.
Forma si distributia vacuolelor in celula variaza in functie de starea fiziologica a celulei, de varsta ei, de tipul celular si de actiunea diferitilor factori de mediu. Asa de exemplu, referindu-ne la actiunea factorilor de mediu, daca unele celule epidermice cu o vacuola mare sunt intepate de anumite insecte, vacuola se imparte in mai multe vacuole mici.
Rolul fiziologic al vacuolelor implica:
▪ Depozitarea substantelor osmotic active (ioni, glucide solubile, acizi organici etc), care genereaza presiunea osmotica si potentialul de membrana.
▪ Prin modificarea turgescentei vacuolei se determina miscarile de inchidere si deschidere a stomatelor, precum si miscarile unor organe ale plantelor (nastii).
Odata cu aparitia filogenetica a vacuolelor s-au format si mecanisme ce regleaza si determina turgescenta. Aparitia fenomenului de turgescenta a constituit o achizitie importanta in evolutia plantelor, conferind indivizilor vegetali o rigiditate mai mare, necesara in actiunea lor de cucerire a uscatului. Aparitia vacuolei face posibila crearea in interiorul celulei vegetale a unui mediu apos cu o compozitie specifica, deosebita de cea a mediului extern.
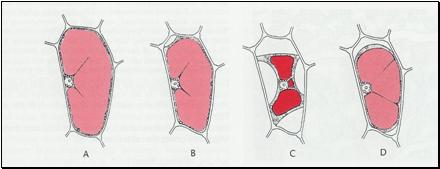
Concentratia molara corespunzatoare acestui suc, procesul de semipermeabilitate a tonoplastului, inconjurarea vacuolei de un strat de citoplasma, cat si prezenta plasmalemei, toate acestea permit vacuolei sa functioneze ca un osmometru, ce joaca un rol important in turgescenta si in stabilitatea celulei (fig. 14).
|
|
|
Fig. 14 - Reprezentarea schematica a gradului de aprovizionare cu apa a celulei vegetale: A - celula turgescenta; B - stadiul de plasmoliza incipienta; C - stadiul de plasmoliza avansata (concava); D - stadiul de deplasmoliza (d. Strasburgher, 1998) |
Este un constituent neviu, care reprezinta un caracter esential de diferentiere a celulei vegetale de celula animala.
Majoritatea celulelor vegetale sunt acoperite de un perete celulozic rezistent, captusit la interior de plasmalema (vezi fig. 3, 8). In regnul vegetal sunt putine celule lipsite de un perete celulozic, celulele numindu-se in acest caz "nude", cum sunt zoosporii si gametii (celulele reproducatoare de la plantele inferioare).
Peretele celular este un constituent solid, relativ rigid, cu o elasticitate limitata. Totalitatea peretilor celulozici dintr-un tesut poate fi imaginata ca o retea densa de lamele sau fibre ce alcatuiesc un schelet al tesutului respectiv, al organului din care face parte tesutul sau chiar al intregului organism.
Peretele celular este permeabil pentru apa si se imbiba cu aceasta, permitand intrarea apei si a diferitelor substante macro- si micromoleculare in celula, precum si iesirea apei din celula, fenomene implicate in procesul de plasmoliza.
Principala lui functie este corelata cu rigiditatea sa relativa si consta in aportul mecanic, de sustinere a formei celulei, din acest motiv numindu-se perete scheletic. Peretele celular are si rol de protectie a protoplasmei. El permite cresterea celulei, precum si schimbul de gaze si de substante nutritive cu celulele invecinate. El permite, de asemenea, intrarea si iesirea apei din celula, prin spatiile dintre microfibrilele celulozice, care constituie apoplastul celular.
|
Intrebari: . Care sunt functiile peretelui celular si cum se mai numeste acesta la celula vegetala? (motivati denumirea). . Enumerati organitele citoplasmatice vii si, separat, organitele ciptoplasmatice nevii din structura celulei vegetale. |
|
Tema: Prezentati succint, pe baza bibliografiei propuse, principalele caracteristici functionale ale celueli vegetale. |
|
Politica de confidentialitate |
| Copyright ©
2025 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |