
Date privind entomofauna nocturna din nord-estul Parcului National Muntii Macinului - Lepidoptera
Caracterizarea generala a muntilor Mǎcin
Asezare geograficǎ
Horstul dobrogean acupǎ coltul de la N-V al judetului Tulcea, dominand Balta Brailei cu 300-400 m si se prelungesc sub forma unei culmi inguste deluroase pana in apropiere de Galati - pintenul Buceagului.
Spre S -V sunt separate de Podisul Babadag prin pasul Iala sau Carapelit.Bine delimitat la V de Balta Brailei spre care cad in trepte, ei depasesc la S-E valea largǎ a Taitei din cursul superior si ajung la Valea Lozovei ingloband astfel douǎ mici culmi.Acest fapt nu e impus atat morfologic cat de linia de incalcare tectonica (falia inversǎ) Luncavita - Consul care-i separǎ de Delurile Niculitel.Dacǎ forma unui triunghi ascutit,muntii Mǎcinului au cea mai mare latime intre Turcoia si Valea Lozovei unde ating 24km.
Ei se desfasoarǎ pe directia NV-SE sub forma unor culmi paralele din ce in ce mai numeroase si mai fragmentate in partea de S si SV, avand o culme principalǎ incepand din pintenul Buceagului pana la Dealul Carapelit.Lantul muntos principal culmineaza cu varful tutuiatu (467m) cea mai mare inaltime din intreaga Dobroge.
Formatiunile geologice predominante aici sant si granitele gnaisice,s isturile, conglomeratele, granitele si dioritele, cuartitele, filitele.Urmeazǎ spre V prima culme secundarǎ reprezentata prin Culmea Cheii cu varful Pricopan (370m) si masivul Megina, cu numeroase varfuri ascutite. Sub raport petrografic in aceasta culme secundarǎ, predominante sunt granitele gnaisice, micasisturile, cuartitele si amfibditele intens dezagregate.
Spre V urmeazǎ o a doua culme secundarǎ, paralelǎ cu cea precedentǎ, formatǎ din mai multe masive mamelonare, cu caracter insular, inecata in loess,constituite din cuartite,micasisturi,filite si dezvoltate in plinǎ zona stepicǎ.In general muntii Mǎcinului din punct de vedere tectonic sub forma unui mare anticlinoriu din care eroziunea foarte accentuate si indelungatǎ a scos la zi granitele cu aspectul lor structural laccolitic.
In prezent ei au aspect miniform, cu grohotisuri pe versanti si depuneri groase de 20-30mm de loess de varsta cuaternarǎ.
CLIMA
Muntii Mǎcinului au o clima specificǎ temperat continentalǎ cu contraste termice de la varǎ la iarnǎ , ploi rare cu caracter de averse, fenomene intense de viscol si precipitatii de 400mm annual.Ca sursǎ energeticǎ principalǎ de care este legatǎ dezvoltarea tuturor proceselor din naturǎ ,radiatia solarǎ prezintǎ aici cele mai ridicate valori din tara peste 125kcal/cm2.
Regimul termic al aerului
Izoterma de 10 grade Celsius este cea care contureazǎ cele doua masive muntoase deluroase.
Repartittia si regimul norilor
Numǎrul mai mare al zilelor acoperite de nori este de 140 de zile iar al zilelor senine ajunge panǎ la 110 zile pe an.
Precipitatiile atmosferice
Sunt destul de scǎzute, iar secetele destul de frecvente ca urmare a deficitului de umiditate.Cantitatea medie multianualǎ atinge in zona inalta a muntiilor Mǎcin 500 mm chiar 600 mm.Intervalul de timp in care nu cad precipitatiile au o duratǎ foarte mare, dar totodatǎ cantitatea maximǎ cǎzute in 24 ore si ploi torentiale pot depasi cu peste 1700% cantitatea medie lunarǎ si cu peste 170% pe cea anualǎ.
Aceasta aratǎ caracterul continental excesiv regional.Caracterul torential al ploilor provoacǎ eroziuni torentiale accentuate,in special in depozitele loessoide.
Presiunea atmosfericǎ,vanturile si regimul lor
Presiunea atmosfericǎ are in general valori lunare si anuale ce depasesc 1000mbarr, atingand 1019 mbarr in luna ianuarie si 1011 mbarr in luna iulie, media anualǎ fiind de 1016 mbarr.Desele invazii de aer continental din timpul iernii sant insotite de cresterii ale presiunii atmosferice la nivelul de 1020 mbarr.
Regimul vanturilor e strans legat de circulatia generalǎ atmosfericǎ de pe teritoriul tarii determinate la rǎndul ei de activitatea marilor centre de actiune atmosfericǎ si de conditiile locale.In muntii Mǎcinului predominǎ vanturiile de N-E si E cu o viteza ce trece de 2 m/s.
Analizate pe anotimpuri, vanturile de N si N-V sau S-E primǎvara reprezinta sezonul cel mai agitate iar vara este cel mai linistit.
Hidrografie
Ea prezintǎ o serie de particularitati cum sant: predominarea raurilor si a torentilor in defavoarea lacurilor si prezenta slabǎ a apelor freatice in urma strans legate de cuvertura de depozite, eluviale, dealuvio-proluviale, leossoide care imbraca atat varfurile cat si versantii sau fundutrile de vǎi largi.Aceste rauri prezintǎ un regim propriu cu scurgere minimǎ de iarnǎ, cu cresteri mari uneori catastrofale de primavarǎ si iarnǎ.
Turbiditatea apei din rauri este accentuatǎ datoritǎ n primul rand loessului ca rocǎ foarte friabilǎ.
Soluri
Privit in ansamblu,invelisul de sol al muntilor Mǎcin reflecta in distributia si dezvoltarea lui, raportul dintre regiunile inalte si joase, iar pe verticalǎ, influenta altitudinii reliefului, conditiile de rocǎ, vegetatia,apa freaticǎ.In regiunile inalte rǎspandirea solurilor este mult mai concentrica.Principalele tipuri de soluri in regiunile inalte sant:cernoziomurile,solurile brune, solurile Silvestre si solurile brune de pǎdure.
Cernoziomul cu tipurile lui (carbonatic,castaniu si levigat) reprezintǎ solulul predominant care impreunǎ cu solurile bǎlane se intlnesc de obicei la periferia muntilor,fiind dezvoltate de obicei pe loess.Solurile bǎlane sunt cele mai tinere soluri dezvoltate pe loesuri.Acestea apar pe pante line si la altitudini absolute panǎ la 150m.
Solurile Silvestre cenusiu inchis sant prezente sub pǎduri quercinee cu elemente mai xerofite.Solurile Silvestre podzolite cenusiu sib run-cenusiu se dezvoltǎ pe reliefulmai ridicat.Solurile Silvestre brune si brune gǎlbui unele slab podzolite se intalnesc pe pantele nordice si nord-estice ale muntilor Mǎcin datorita umiditatii mai mari.
Energia mare de relief, specificǎ regiunii muntoase si deluroase, roca friabilǎ si vegetatie lemnosǎ destul de rarǎ,fac sa aparǎ pe pantele mai accentuate din culmea Pricopanului, portiune cu eroziune accentuatǎ.
Vegetatia
Complexul conditiilor naturale, actiuni modificatoare transformatoare a factorilor antropici si zoogenic(defrisare,pǎsunatul excesiv)au determinat existenta a numeroase formatiunii si asociatii ce au modificat foarte mult aspectul natural al acestui colt dobrogean.Muntii Mǎcinului sant unici si reprezentativi la nivel national datorita originalei combinatii intre caracterul geologic si specificitatea biogeograficǎ atribuite interferentei in aceastǎ zonǎ a limitei sudice central europene a speciilor caucaziene,a limitei nordice a speciilor mediteraneene,balcanice si pontice ca si a limitei de vest a unor specii asiatice.
De asemenea, ei reprezintǎ limita nordicǎ a Peninsulei Balcanice a zonei submediteraneene in general si beneficiind `in acelasi timp de umiditate distincta floristice macedo-tracicǎ.Cele trei zone biogeografice care apar sant:stepa, silvostepa si pǎdurea.Zona de stepǎ se intinde tentacular in depresiunile dintre formatiunile inalte urcand in altitudine panǎ la 50-100m.
Stepa cu graminee adaptate regiunii uscate apare in amestec cu pajisti xerofite stepice sau derivate dominate de:paius (Festuca sulcata,F.vaginata) Negara (Stipa capillata) colilia (Stipa stenoph yla si S.joannis) pe langǎ care apar elemente pontice:otratelul (Onosma avenarium) garoafa (Dianthus palleus) elemente mediteraneene:aison (Apshodeline lutea) si elemente ponto -sud-siberiene:cǎciula mocanului (Rindera umbellate) si mǎrgica (Melica ciliata).
In zona de stepǎ vegetatia lemnoasǎ este reprezentatǎ prin arbusti :pǎducel (Crataegus monogyna) porumbar (Prunus spinosa) si specii caracteristice Dobrogei: paliurul (Paliurus spina-christi) si iasomie (Jasminum fruticans).
Silvostepa
Se interpune ca o zonǎ de tecere de la stepǎ la padure sub forma unor fasii inguste care ajung panǎ la altitudini de 200 m.Ea apare ca o imbinare cu pajistile stepice secundare degradate precum si mici insule de pǎdure. Invelisul ierbos este format din:bǎrboasǎ (Andropogoni ischaemum),firuta (Poa bulbosa) si alte sp xeromezofile.
Palcurile de arbori sant alcǎtuite
din stejar pufos (Quercus pubescens)
si brumǎriu (Quercus
pedunculiflora),
Conditiile de vegetatie din pǎduri permit o mare dezvoltare a arbustilor (pǎducel, corn) si a unui invelis ierbos format din diferite specii, intre care predominǎ gramineele.Tot aici trebuie amintita vegetatia si fauna azonalǎ a pajistilor de vǎi si lunci cu soluri,navene, vegetatia si fauna de balti, pe terenuri cu exces de umiditate, vegetatia si fauna de sǎrǎturǎ , pe terenuri joase cu exces de sǎruri in sol si apǎ ca si vegetatia de nisipuri cu exces de uscǎciune (zona dunelor dintre Jijila si Vacǎreni) care in general ocupǎ suprafete foarte reduse.
Fauna
Vertebratele din zonele de stepǎ si silvostepǎ sǎnt numeroase mai ales rozǎtoare:popandǎul (Citellus citellus),soarecele de camp (Microtus arvalis), grivanul dobrogean (Mesocricetus newtoni),broastele de pǎmant (Pelobates fuscus), si broastele raioase (Bufo bufo).Pe pantele insorite se intalneste destul de frecvent broasca testoasǎ de uscat (Testudo greca ibera) specificǎ Dobrogei.
Dintre reptilele caracteristice acestei zone sant:gusterul vǎrgat (Lacerta trilineata dobrogica), soparla de pustiu (Eremias arguta deserti), vipera cu corn dobrogeanǎ (Vipera ammodites montandoni).
Pǎsǎrile sant destul de numeroase unele populeazǎ frunzarul arborilor- pitigoiul mare (Parus major),pitigouil mic (P.coeruleus), silvia mica (Sylvia corruca).
Dintre mamifere mai frevente sant:cǎprioara (Capreolus capreolus), mistretul,lupul (Canis lupus).
Insecte; entomofauna zonei este foarte bogatǎ in specii, fiind studiatǎ de specialisti inca din anul 1865 (Mann, 1866). Numǎrul total de taxoni de pe teritoriul parcului national Muntii Mǎcinului nu este cunoscut cu precizie, insa totalul depaseste cu siguranta cifra de 5000 de specii. Pentru a ne face o imagine despre entomofauna zonei, doar din lista 941 de specii de lepidoptere citate in 2000 din nordul Dobrogei (Rakosky, Wieser, 2000), peste 90% au fost mentionate de la Greci, Pricopan, Turcoaia, Horia.
Incadrarea generalǎ a muntilor Mǎcin
Muntii Mǎcin sunt situati in sud-estul Romaniei, respectiv in nord-vestul judetului incadrati intre 28°07' longitudine esticǎ, respectiv 45°01' si 45°21. Altitudinal acestia se incadreazǎ intre 7 si 467 m, reprezentand zona cea mai Dobrogean, fiind situati in apropierea limitelor vestice ale Rezervatiei Biosferei.
Situati intr-un climat moderat-continental ei sunt caracterizati prin temperaturi intre 10,8°C si 9°C, respectiv precipitatii intre 480,4 mm si 600 mm, fiind cei din Romania.
Localizarea si incadrarea teritorial administrativ
Parcul national este situat pe teritoriul administrativ al orasului Mǎcin si comuna Cerna in vestul parcului, comuna Cerna amplasatǎ in vestul si sud-vestul parcului Hamcearca si Luncavita-situate in estul acestuia precum in Jijila in partea dreaptǎ suprafata totalǎ de 11321 ha a parcului, 30 ha apartin Consiliului local al orasului Mǎcin ocupand o suprafatǎ de 11291 ha apartinand Directiei Silvice Tulcea, respectiv Ocoalele Silvice Mǎcin.
Vecinatati si limite
In general limita parcului este constituitǎ prin limita de contact intre fondul formatiei si terenurile agricole sau pasunile apartinand localitǎtilor in jur.Astfel, in partea de vest parcul se invecineazǎ cu terenurile agricole si pasunile orasului Mǎcin si comunei Greci, in vest si in sud-vest cu cele ale comunei Cerna, comunelor Hamcearca si Luncavita, iar la nord cu cele ale comunei Jijila.Limita in teren pe cea mai mare parte a perimetrului prin borne silvice, cu exceptia Raioasa unde acestea sunt delimitate doar prin drumuri de pǎmant.
Suprafatǎ si zonare
Parcul National Muntii Mǎcin se intinde pe o suprafata de 11.321 ha. Din care reprezintǎ zonele de conservare special desemnate conform Ordinului MAPA, privind aprobarea zonǎrii interioare a parcurilor nationale.
Parcul National Muntii Macin
Muntii Mǎcin reprezintǎ o componentǎ cheie mostenirii naturale a tarii si a lumii intregi.Parcul National Muntii Mǎcinului are o suprafata totalǎ de 11.321 hectare, fiind structurat in trei perimetre:Culmea Pricopanului, Culmea Mǎcinului si Culmea Luncavita, in cadrul acesteia din urmǎ evidentiindu-se Rezervatia Valea Fagilor (154 ha).
Rezervatia Valea Fagilor, inclusǎ in parc, este o insula relictarǎ de fagi ascunsǎ intr-o vale ingustǎ si umedǎ, cu exemplare ce ajung la 1 m grosime si 38 m inaltime. In compozitia pǎdurii existǎ ospecie unicǎ- fagul tauric (Fagus taurica) provenitǎ din incrucisarea fagului oriental cu carpenul si teiul argintiu.Fǎgetul din Rezervatia Valea Fagilor- Luncavita este considerat relict tertiar, fiind un tip de pǎdure unic in tara.
Caracterizarea morfologicǎ a Muntilor Mǎcin
Ca morfologie, Dobrogea de Nord prezintǎ relief tipic de peneplena, avand caracteristic Muntii Mǎcinului, erodati, rǎmasi ca un rudiment al sistemului cutat hercinic. Cota maximǎ este 458 m in masivul Greci.
Reteaua hidrograficǎ
este foarte saracǎ.Vǎile sunt foarte largi, unele avand numai temporar apǎ.Produsele de eroziune sunt transportate foarte putin panǎ la baza pantei si sunt numai partial antrenate in lungul unor vǎi cu apǎ.Vǎile largi sunt acoperite cu o cuvertura groasǎ de depozite loessoide si deluvial-proluviale, care mascheazǎ depozitele mai vechi. Din aceasta cauzǎ, deschiderile se gǎsesc numai pe crestele dealurilor sau in lungul versantilor abrupti.
Subunitatea Muntilor Mǎcin (zona Mǎcin) este situatǎ in partea de NV a orogenului nord-dobrogean si are o latime de 15 km intre Dunǎre la Vest si o linie de dislocatie spre Est , care uneste localitatea Luncavita de pe Dunaǎre si dealul Consu; de pe valea Taitei, denumita dislocatia Luncavita-Consul.
La vest de Dunǎre, datele de geofizicǎ si forajele au conturat prelungirea Orogenului nord-dobrogean afundat.Spre SE, zona Mǎcin se prelungeste pe sub bazinul Babadag.Cea mai mare parte din zona Mǎcin este ocupatǎ de formatiuni sedimentare de varsta paleozoicǎ, al cǎror substrat este format din sisturi cristaline.
Orogeneza hercinicǎ a dus la cutarea si ridicarea zonei Mǎcin.Ulterior, zona Mǎcin nu a mai fost afectatǎ in mod sensibil de miscari cu efect plicativ, ci a functionat ca sistem cutat emers in tot timpul Permianului, iar in Triasic au fost acoperite de ape numai partile marginale.Doar miscarile din faza chimericǎ veche au reactivat o fracturǎ preexistentǎ pe directia Luncavita-Consul, in lungul cǎreia zona Mǎcin are tendinta de incǎlecare peste zona Tulcea, prinzand sub planul de falie depozitele Triasicului superior.
Se poate concluziona cǎ structura majorǎ a zonei Mǎcin este efectul orogenezei hercinice.Exploatǎrile de cariere, prin activitatile specifice desfasurate (metode de exploatare cu explozivi, statii de concasare-sfǎramare-sortare, activitati de transport cu utilaje grele) au condus la disparitia faunei din apropierea carierelor, la deteriorarea vegetatiei in zonele afeectate de exploatǎri si au produs un impact deosebit, negativ asupra peisajului, prin acele "gǎuri negre", scobituri ireversibile in scoarta terestrǎ.
Incǎ de la promovarea ideii de arie protejatǎ si infiintarea acesteia prin aprobarea Consiliului Judetean Tulcea si a autoritatilor judetene si locale, s-a pus problema si s-a trecut treptat la inchiderea carierelor existente in interiorul ariei protejate.Singurele cariere rǎmase in activitate sunt cele douǎ aflate in vecinatatea Parcului,dar aflate la distante apreciabile fata de acesta, si a cǎror activitate s-a considerat a nu afecta in mod deosebit ecosistemele din zonǎ.
Potentialulul turistic al Muntilor Mǎcin
Elementele care exprimǎ potentialul turistic al Muntilor Mǎcin, pot constitui factori determinanti pentru dezvoltarea turismului in cadrul acestei frumoase zone a Romaniei.
Obiectivele turistice, usor accesibile de pe traseele marcate sau de pe drumuri, sunt formate din elemente ale potentialului turistic natural (peisaj, rezervatii naturale, monumente ale naturii), precum si de elemente ale potentialului turistic antropic (monumente istorice si de arhitecturǎ, obiective culturale si de artǎ popularǎ).
Potentialul turistic natural este reprezentatǎ prin urmatoarele obiective:Varful Tutuiatu (467 m); Varful Capusa (433 m); Varful Moroianu (434 m); Varful Pricopcea (410 m) ; Varful Sulucu Mare (370 m) si Culmea Pricopanului; Punct fosilifer Dealul Bujoarele rezervatie naturalǎ geologicǎ; Pǎdurea Valea Fagilor (rezervatie naturalǎ botanicǎ ) Lacul Sǎrat; Broasca testoasa dobrogeanǎ (monument al naturii) .
Descrierea ariei protejate
Muntii Mǎcin, rezultat al orogenezei hercinice, reprezintǎ cea mai veche formatiune muntoasǎ din tara si printre cele mai vechi din Europa. Localizati in partea de sud-est a Romaniei (in judetul Tulcea), s-au format intre erele Paleozoicǎ si Mezozoicǎ, si au o suprafata totalǎ de 50 000 ha si inaltimea maximǎ de 467 m (Vf. Tutuiatu). Ei reprezintǎ un unicat din punct de vedere floristic, faunistic si peisager, fiind situati la o altitudine joasǎ si intr-o pozitie biogeograficǎ deosebitǎ, ce determinǎ in plan floristic interferenta limitei nordice a speciilor mediteraneene, balcanice si pontice, cu limita sudica a speciilor central europene si caucaziene, si limita vesticǎ a catorva specii Asiatice.
Recunoasterea caracterului reprezentativ si unic la nivel national a Muntilor Mǎcinului precum si a valorii internationale conferite in special prin prezenta numeroaselor specii amenintate cu disparitia, s-a materializat prin constituirea Parcului National Muntii Mǎcinului, in suprafatǎ totalǎ de 11.321 ha, incluzand douǎ lanturi muntoase principale: Pricopan-Megina (in capatul nord-vestic) si Mǎcin (in partea centrala si sud-estica) separate prin depresiunea Greci
Din suprafata totalǎ a parcului, 11.291 ha apartin Administratiei Nationale a Pǎdurilor, din care 10.160 ha sunt pǎduri, 940 ha sunt terenuri neproductive, 130 ha sant habitate stancoase si 61 ha sunt alocate administratiei, restul de 30 ha fiind pǎsune comunalǎ ce apartine Consiliului Local Mǎcin.
Unul din principalele argumente pentru infintarea acestui parc este valoarea ecologicǎ remarcabilǎ a acestor munti si prezenta multor specii floristice care sunt periclitate atat la nivel national cat si international. In aceastǎ regiune existǎ aproximativ 1900 specii de plante reprezentand peste 19% din flora Europeanǎ, numǎrul acestora fiind comparabil cu flora bogatǎ a insulelor Creta si Corsica. Mǎcin si imprejurimile lor sant singurele zone din Romania unde inca mai existǎ suprafete importante de vegetatie naturalǎ de stepǎ care nu se gaseste in alte parti ale Romaniei sau altundeva in Balcani, Parcul National Muntii Mǎcin reprezentand singurul loc din tara care protejeazǎ acest tip de vegetatie. Sant incluse aici 27 de specii si subspecii de plante endemice (Campanula romanica, Corydalis solida ssp slivenensis, Euphorbia nicaeensis ssp cadrilateri, Moehringia grisebachii, M. Jankae, Silene cserei).
Lista faunei Muntilor Mǎcin include in jur de 2000 de specii de nevertebrate (aproximativ 100 de specii de fluturi au fost reconfirmate in anul 2000, ca fiind prezente), 7 specii de amfibieni, 11 specii de reptile (incluzand specii rare , incluse in anexele Directivei Habitate si ale Conventiei de la Berna:Testudo graeca, Ablepharus kitaibelli, Lacerta trilineata, Elaphe quatorlineata, Viper ammodytes), si cel putin 187 de specii de pǎsǎri (specii rare incluse in anexele Directivei PASARI:Anser erythropis, Aquila clanga, Aquila pomarina, Crex crex, Hieraetus fasciatus, Pernis apivorus, Neophron pernopteris, etc), si 40 de specii de mamifere (incluzand specii adaptate la stepǎ ca Spermophilus citellus- inclusǎ in Anexa 2 a Directivei Habitate si Coventia de la Berna, Vormela pergusuna,Canis aureus etc).
Pentru pǎsaǎrile rǎpitoare din Romania Muntii Mǎcin reprezintǎ cea mai importantǎ zonǎ de cuibarit , fiind de asemeni un important loc de pasaj pentru cele migratoare.
O parte din insectele gǎsite in Muntii Mǎcin sunt noi pentru stiinta, iar speciile Calimorpha quadripunctaria si Morimus funereus sunT incluse in Anexa 2 a Directivei Habitate si in Conventia de la Berna.
Principalele tipuri de habitate prezente in Muntii Mǎcinului sunt: pajistile pietroase, calcaroase sau bazofile cu Alysso-Sedion albi, pǎdurile de Tilio-Acerion pe povarnisuri, grohotisuri si viroage, pantele de rocǎ silicioasǎ cu vegetatie casmofiticǎ, pǎdurile de fag Luzulo-Fagetum, pǎdurile termofile de Fraxinus angustifolia si pǎdurile de stepǎ euro-siberianǎ cu Quercus sp (incluse in Anexa 1 a Directivei Habitate) precum si: stepe continentale, pǎduri de fag, pǎduri de stejar si carpen si pǎduri termofile si supra-mediteraneene de stejar (incluse in Conventia de la Berna). Dintre acestea numai habitatele de stepǎ sunt pasunate moderat, restul fiind bine conservate.
Aria protejatǎ, Muntii Mǎcin este situatǎ inr-una dintre dintre cele mai remarcabile regiuni ale Europei, si poate fi consideratǎ ca o componentǎ cheie a mostenirii naturale a Romaniei si a intregii lumi. Bogatia biodiversitatii sale, mǎrimea sa si amplasarea, dau acesteia posibilitatea de a deveni una dintre cele mai importante arii protejate din regiune. In orice caz planurile pentru statutul ariei protejate trebuie sa fie in concordanta cu necesitatile si asteptarile comunitatii locale care nu poate fi separatǎ de natura Muntilor Mǎcin. Prezenta acestora, interesul si activitatile lor au o legaturǎ importantǎ si un potential impact asupra succesului si sustinerii oricarea arii protejate sau administrate din regiune.
Romania are una din pǎdurile temperate cele mai importante din punct de vedere biologic din lume. Muntii Mǎcin sunt unii dintre cei mai vechi munti din Romania. Unicitatea lor este rezultatul combinatiei dintre caracterul geologic intrinsec si specificitatea si locttia lor biogeograficǎ. Acesti munti prezintǎ ecosisteme caracteristice pentru stepa ponto-balcanicǎ, pǎduri sub-submediteraneene si balcanice, impreunǎ cu un numǎr mare de specii periclitate.
Habitatele biogeografice si comunitatile din muntii Mǎcinului sunt complet diferite de cele din muntii Carpati (unde se aflǎ deja in derulare initiative de protejare a ecosistemelor specifice in cadrul altor Parcuri Nationale romanesti). Din punct de vedere geografic muntii Mǎcinului se situeazǎ la o altitudine joasǎ intr-o pozitie unicǎ din punct de vedere biogeografic.
Din punct de vedere floristic ei reprezintǎ limita nordicǎ a speciilor mediteraneene, balcanice si pontice, limita sudicǎ a speciilor central europene si caucaziene, si limita vesticǎ a catorva specii asiatice.
Toate comunitǎtile de plante din zonǎ sunt considerate rare la nivel european.Anumite asociatii floristice formeaza comunitati unice care se intalnesc numai in aceastǎ parte a lumii (de ex: Asociatia Gymnospermo-Celtetum). Acesti munti conservǎ comunitati de plante si specii reprezentative sau periclitate, multe dintre ele endemice, pentru unitatea biogeograficǎ a Platoului Dobrogean. Numǎrul speciilor de plante variazǎ in aceastǎ regiune intre 1779 si 1911 functie de referinte.Ele reprezintǎ peste 19% din flora europeanǎ si sant comparabile cu flora bogatǎ a insulelor Creta si Corsica.
Unul dintre argumentele cele mai importante pentru infiintarea acestui Parc, si totodata o remarcabilǎ parte a valorii ecologige a acestor muntii,o reprezintǎ prezenta numǎrului mare de specii amenintate de flora (>72) de statut national si international. 1,4% sunt considerate acum "extincte", 17% "vulnerabile" sau "rare" (conform Listei Rosii). Un exemplu al importantei acestei arii la nivel de comunitati ecosistemice, il reprezintǎ pǎdurea de fag si carpen dobrogean, care este unicǎ in Romania. Un alt exemplu, la nivel de specie, este cel al lui Campanula romanica (clopotelul dobrogean), Parcul Muntii Mǎcinului fiind singurul Parc din lume care oferǎ protectie acestei specii endemice, intalnita in aceastǎ regiune. In total aceastǎ arie detine 27 de specii de plante endemice. In orice caz se pare cǎ datele existente sunt putine si cǎ ar putea exista mai multe specii decat cele inregistrate panǎ in prezent.Dobrogea este singura regiune din Romania unde inca mai existǎ suprafete importante de vegetatie stepicǎ, unice in Romania si in Balcani. Muntii Mǎcinului reprezintǎ singurul Parc National din tarǎ care protejeazǎ acest tip de vegetatie, foarte rarǎ acum in Europa. Tocmai de aceea acest parc ar fi singurul din lume care conservǎ habitate specifice, comunitati de plante, specii endemice, subendemice si relicte ale platoului dobrogean, o arie biogeograficǎ de importanta internationalǎ, recunoscutǎ.
In ciuda naturii unice biogeografice a Muntilor Mǎcinului, cu atractiile lor speciale florale si faunistice, informatiile asupra diversitatii faunistice sunt putine. Dintre cele 187 de specii de pǎsǎri observate in zona parcului, 60% |(aprox. 112 specii), sunt considerate "Vulnerabile", "Rare", sau "Posibil extincte", conform criteriilor IUCN, si multe dintre celelalte specii sunt considerate "Insuficient cunoscute". Muntii Mǎcinului reprezinta cea mai importantǎ zonǎ de cuibarit din Dobrogea pentru pǎsǎrile de pradǎ si de asemeni reprezintǎ o importantǎ zonǎ de pasaj pentru cele migratoare.
Dintre cele 41 de specii de mamifere inregistrate aici, 11 sunt protejate prin conventii internationale. Existǎ peste 979 de specii de lepidoptere inregistrate, iar cercetǎrile de entomologie recente realizate in Muntii Mǎcinului au scos la ivealǎ inca 14 specii noi pentru Romania si 3 specii pentru stiinta.
Existǎ 7 specii strict protejate de amfibieni iar toate cele 11 specii de reptile inregistrate in zona Muntilor Mǎcinului sunt de asemeni protejate prin Conventia de la Berna. Aceste comunitati si specii nu se gǎsesc in cadrul altor parcuri sau arii protejate din Europa sau imprejurimi.Fǎrǎ acest parc, protejarea biodiversitatii internationale ar fi incompletǎ.Valoarea internationala a biodiversitatii ecosistemelor din Muntii Mǎcinului este incontestabilǎ.
Istoricul cercetarilor asupra lepidopterelor in nord-vestul Dobrogei
Pentru entomofauna Dobrogei de Nord, punctul de referinta pentru o serie de grupe de insecte il constituie lista publicata in anul 1866 de Joseph Mann care in anul precedent efectuase o campanie de colectare in nordul Dobrogei, materialul intrand ulterior in patrimoniul Muzeului de Stiinte Naturale din Viena. Lista sistematica, ce cuprinde si o serie de informatii ecologice privind diveresele specii, include lepidoptere, coleoptere, ortoptere, neuroptere, diptere, heteroptere, homoptere si arahnide.
Lepidopterele identificate reprezinta o parte insemnata a materialului colectat, fiind mentionati peste 900 de taxoni. Aria acoperita in timpul deplasarilor pe teren este reprezentata atit de zonele impadurite din Podisul Babadagului cat si de silvostepa si de Lunca Dunarii. Chiar daca Mann nu a colectat de pe teritoriul actual al parcului national, totusi datele oferite sunt extreme de valoroase, dat fiind faptul ca zona respectiva era in perioada sfarsitului secolului XIX impadurita aproape uniform si entomofauna nu prezenta diferente notabile. Din acest motiv avem toate motivele sa credem ca ceea ce Mann colectase in zonele estice ale nordului Dobrogei nu diferea de ce se gasea in vestul regiunii.
In perioada 1973 - 1975, A. Popescu Gorj efectueaza o serie de colectari in nordul Dobrogei, relevind prezenta in aceasta regiune nu numai a unor specii care nu mai fusesera citate din 1866 dar si a unor specii noi pentru entomofauna Dobrogei.
In perioada de dupa 1990, cercetari sistematice sunt efectuate in zona mai ales pentru lepidoptere de catre L. Rakosy, C. Wieser s.a. iar mai recent de catre colectivul de la Institutul de Cercetare Dezvoltare "Delta Dunarii" din Tulcea sau Muzeul de Istorie Naturala "Grigore Antipa" din Bucuresti (E.Pisica, I.S. Iorgu - Orthoptere, M.Stan - coleoptere stafilinide).
Metodǎ si material de lucru
Pentru fauna de lepidoptere s-au efectuat colectari calitative pe durata a cinci luni in special in zona cantonului silvic Cetatuia. In acest punct a fost amplasata o pereche de capcane luminoase care au functionat pe timpul noptii in perioadele in care s-au efectuat deplasari pe teren. Pentru investigarea entomofaunei nocturne, s-au efectuat colectari la surse de lumina artificiala in zona cantonului silvic Cetatuia. Capcanele luminoase clasice, au functionat in toate perioadele in care au fost efectuate deplasari si colectarile de lepidoptere au fost mai aes calitative. Pe de alta parte, materialul biologic colectat a fost conservat in intregime pentru toate grupele taxonomice, in acest fel realizandu-se si o baza de date pentru entomofauna nocturna. Lepidopterele au fost conservate in stare uscata - plicuri entomologice. O parte din exemplare au fost etalate imediat sau dupa umezire.

Materiale necesare: Pentu colectarea insectelor nocturne sant necesare:o sursǎ de luminǎ,fileu entomologic, substante anesteizante (eter etilic,alcool etilic de 70 grade) cateva borcane cu dop si hartie de calc.
Metode de colectare: Pentru colectarea insectelor nocturne este necesar o sursǎ de luminǎ si o sticlǎ sectionatǎ in jumatate ce se prinde de sursa luminoasǎ iar insectele in zbor cad in capcanǎ luminoasǎ ,iar pentru prinderea fluturilor a fost necesar utilizarea fileului entomologic.
Metode de conservare: Insectele colectate au fost transportate la domiciliu cu substante toxice apoi am inceput trierea lor.
Prelucrarea datelor: Pentru identificarea cu precizie a insectelor conservate am folosit lupa binocular si le-am comparat cu cele din literature de specialitate.
Identificarea materialului: Identificarea materialului a fost efectuata atat pe teren cat si in conditii de laborator utilizand determinatoare de specialitate sau lucrari monografice.



Rezultate si discutii
Au fost efectuate colectari cantitative - incercandu-se sa se surprinda intregul spectru taxonomic al insectelor nocturne, cat si colectari calitative pentru lepidoptere. Dintre familiile de lepidoptere nocturne, s-a insistat in special asupra geometridelor si noctuidelor - grupe taxonomice importante, cu un mare numar de specii in zona. In afara acestor familii, materialul colectat a inclus specii din familii ca Drepanidae, Axiidae, Syntomidae, Arctiidae, Notodontidae, Sphingidae, etc. Microlepidopterele nu au putut fi analizate pana la specie.

Prezentarea grupelor taxonomice din probe
Ordinul Hymenoptera
Din acest ordin fac parte insecte ca albinele, viespile, furnicile, bondarii si multe alte grupe de himenoptere libere sau parazite.
Capul este de tip ortogonat.Formele aripate au totdeauna bine dezvoltati ochii cu fatete,iar ocelii sant prezenti.In ceea ce priveste antenele, ele au o alcǎtuire foarte diferitǎ la diferitele grupe de himenoptere, dar cel mai adesea sant antene filiforme.
Segmentele toracelui sant de regulǎ concrescute, cel putin pe partea dorsalǎ si numai in regiunea sternalǎ se disting bine segmentele, ca si partile laterale.Protoracele si metatoracele sant intotdeauna mai mici decat mezotoracele, care ia cea mai mare dezvoltare.Protoracele este de regulǎ foarte ingust la partea dorsalǎ si se lateste numai pe partile laterale.
Natura membranoasǎ a aripilor nu diferǎ de aceea a aripilor membranoase alte altor insecte.Caracteristic este insa sistemul de angrenare a celor douǎ aripi prin frana formatǎ din carligele de la partea anterioarǎ a aripii posterioare si indoitura de la partea posterioarǎ a aripii anterioare.
Nervatia este caracteristicǎ diferitelor familii , atat in nervuri cat si in celulele formate de aceste nervuri.Aceastǎ nervatie este mult specializatǎ si adesea greu de interpretat.
Piesele bucale sant formate la multe himenoptere din piese de tip ortopteroid si mult asemǎnator cu cel de la ortoptere, deci cu aparat bucal masticator,insa la formele mult specializate aparatul bucal este transformat pentru masticat si lins, tipul general de aparat bucal rǎmanand tot cel ortopteroid, deoarece mandibulele sant totdeauna prezente.
Picioarele himenopterelor, desi organizate dupǎ tipul general de alcǎtuire, au totusi numeroase particularitati, care sant mult folosite in sistematica acestor insecte.Astfel, trohanterul la multe grupe de himenoptere prezintǎ o gatuiturǎ transversalǎ care dǎ impresia cǎ sant douǎ articole de trohanter.La albine metatarsul picioarelor posterioare este foarte dezvoltat in comparatie cu celelalte articole ale tarsului.La multe grupe picioarele anterioare prezintǎ o particularitate in structura tibiei si a metatarsului care formeaza asa-numitul aparat curatitior al antenei.Acesta constǎ dintr-un dinte lung si puternic la extremitatea distalǎ a tibiei si un rand de peri spiniformi la partea proximalǎ a metatarsului.La partea apicalǎ a tibiilor au spini caracteristici.
Abdomenul este intotdeauna bine delimitat de torace si la multe forme el este pedunculat.De regulǎ ultimele segmente se pot intelescopa.Femelele posedǎ un ovipozitor de tip ortopteroid, compus din trei perechi de teci.La formele sociale ovipozitorul este transformat in ac, pierzandu-si functia de ovipozitor si fiind legat de o glandǎ cu otravǎ.Sant si forme la care ovipozitorul, desi este foarte dezvoltat, totusi el nu este vizibil, fiind retras in pozitie de repaus in interiorul abdomenului.
Dezvoltarea este holometabolǎ; larvele sant apode la himenopterele superioare si numai la unele grupe inferioare sant larve eruciforme polipode.Pupa este de tipul pupǎ liberǎ si cel mai adese ea este inchisǎ intr-un cocon mǎtǎsos.Ingrijirea progeniturii a luat la formele superioare foarte pronuntatǎ.Impuparea si dezvotarea pupei panǎ la forma dea adultǎ au loc in medii foarte variate:in plante, in corpul altor insecte,in cuiburi construite din cearǎ,din celulozǎ,din pǎmant.
Ordinul Homoptera
Numele ordinului derivǎ de la gr.homos=asemenea,acelasi si pteron=aripǎ.Insectele cu ambele perechi de aripi cu aceeasi consistentǎ, fiind aripi membranoase;in repaus ele sant asezate deasupra abdomenului, ca un acoperis de casa, in genere depasind abdomenul in lungime.Aparatul bucal este identic cu cel de la heteroptere, rostrul adaptat pentru intepat si supt.Baza rostrului este apropiatǎ de coxele anterioare, metamorfoza este gradatǎ,afara de unele forme specializate.
Aripile homopterelor sant membranoase,numai la unele forme aripile anterioare fiind pergamentoase.Sant si forme aptere;astfel,femelele de coccide sant lipsite de aripi, masculii de afide sant uneori aripati,alteori apteri,femelele partenogenetice de afide sant si aripate si aptere.Masculii de coccide au o singurǎ pereshe de aripi, cele anterioare,perechea a doua fiind reduse la haltere cu formǎ de mici maciuci.
Nervatia aripilor este mult redusǎ la cele mai multe homoptere.Ca tip de nervatie se considerǎ nervatia de la aripa de Cicada.
Homoperele au fruntea intoarsǎ ventral si indreptatǎ inapoi, capul fiind in intregime atasat la torace in asa mod,incat laturile capului vin in contact cu coxele picioarelor anterioare, rostrul intinzandu-se intre coxele anterioare.
Mandibulele si maxilele sant stiliforme,fiind inchise in tubul sau teaca formatǎ de labium.
Homopterele au ochi fatetati si oceli.Antenele lor sant in genere scurte, iar cand sunt mai lungi sant formate totusi din putine articole.De regulǎ la cap se distinge un clipeus mare,lat,care ocupǎ mare parte din partea anterioarǎ a capului, iar fruntea la multe homoptere lipseste cu totul sau este mult redusǎ.In multe publicatii se confundǎ la homoptere clipeus cu fruntea.
Toracele este normal constituit.Picioarele sant cele mai adesea picioare pentru mers,homonome la unele forme cele posterioare fiind adaptate pentru sǎrit.Formele sedentare au piciarele mult reduse.Tarsele au 1-3 articole.
Abdomenul este legat de torace printr-o regiune latǎ;el nu poartǎ niciodata cercisi se terminǎ printr-o armaturǎ genitalǎ, care prezintǎ particularitati de conformatie la diferitele familii ale ordinului si este diferitǎ la cele douǎ sexe.Armatura genitalǎ are mare importantǎ la determinarea speciilor.Armatura femelǎ este constituitǎ din oviscapt (ovipozitor),care are forma alungitǎ si alcatuitǎ din doua piese care se apropie si este vizibil pe partea ventrala a abdomenului; in unele cazuri,el este mascat de plǎcile genitale.
Homopterele sant heterometabole.Viviparia si reproducerea prin schimb de generatii sant fenomene commune in interiorul acestui ordin de insecte.
Ecologia si biologia lor prezintǎ aspecte variate,totusi toate homopterele sant sugatoare de sucuri vegetale ǎi de regula insecte terestre.Numeroase forme sant dǎunatoare plantelor cultivate si din acest punct de vedere studiul homopterelor este deosebit de important.
Ordinul COLEOPTERA
Coleopterele sant insecte care in marea lor majoritatea au corpul tare, scortos,adesea bombat.Corpul are o o simetrie bilateralǎ, la care cele mai multe forme fiind alungit.Cel mai pregnant caracter anatomic al acestui ordin este prezenta aripilor acoperitoare, tari, scortoase,sclerificate, acoperind la cele mai multe dintre ele intreg abdomenul (elitrele).
In cele mai multe cazuri,elitrele au pierdut total nervatia si posedǎ o structurǎ complicatǎ ,care le asigurǎ rezistenta. In timpul zborului, elitrele se ridicǎ in sus, dar la unele coleoptere nu se ridicǎ (cetoniinde). Acestea lasǎ neacoperit din torace numai pronotul si uneori o mica portiune din mesonot, cu forma triunghiularǎ (Scutellum). La putine coleoptere elitrele sunt scurte si nu acoperǎ intreg abdomenul(Staphylinidae,Meloidae)
Aripile posterioare (perechea a doua),situate dedesuptul elitrelor, sant membranoase si subtiri (alae), mai lungi decat elitrele si stranse dedesuptul acestora, adesea indoite. Nervatia acestor aripi membranoase este caracteristicǎ, fiind consideratǎ ca o nervatie mult specializatǎ.
La multe carabide,cuculionide si alte coleoptere lipsesc aripile membranoase, unele dintre acestea avand chiar elitrele sudate (Lethrus,Blaps,Gnaptor). La putine forme de coleoptere lipsesc chiar ambele perechi de aripi (femelele de Lampyridae).
Capul celor mai multe coleoptere este adesea varat mai mult sau mai putin in protorace si are o pozitie prognatǎ, gura cu piesele bucale fiind indreptata inainte.
Aparatul bucal este de tip masticator, ortopteroid, adaptat pentru tǎiat, ros si mestecat. Dintre piesele bucale, de regulǎ mandibulele sunt foarte bine dezvoltate.
Antenele coleopterelor au infatisǎri foarte felurite, dar numǎrul articolelor nu trece de 10-11.
Protoracele are dorsal forma unui scut si este articulat mobil cu mezotoracele. Mezotoracele este mai mic.
Picioarele sant tipice pentru mers, evident homonome.
Numǎrul articolelor tarsale variazǎ de la 3-5 si se terminǎ intotdeauna cu douǎ gheare. In seria coleopterelor se constatǎ diferite specializari ale picioarelor si implicit, diverse conformatii. La toate coleopterele coxele picioarelor posterioare sant latite si sudate cu toracele, incat par a face parte din scleritele toracelui.
Abdomenul, cel mai adesea alungit si turtit dorso-ventral, se leagǎ de torace printr-o baza latǎ. Primul segment abdominal este sudat cu metatoracele. Nici unul dintre segmentele abdominale nu prezintǎ cerci sau astfel de apendici la adulti. Segmentele terminale alcǎtuiesc oviscaptul, care poate iesi in afara.
Dezvoltarea coleopterelor este tipic holometabola si numai exceptional hipermetabolǎ. Niciodat larvele nu seamanǎ cu adultii;de regulǎ, larvele poartǎ pe segmentul 9 pseuducerci; de obicei larvele au alt fel de viata decat adultii. Larva tipicǎ are aparat bucal masticator si trei perechi de picioare toracice articulate.
Pupa este de tipul pupǎ liberǎ, uneori pupa obtectǎ (Coccinellidae).
Ordinul Diptera
Dipterele (popular mustele, tanǎtarii, tǎunii) sant insecte cu corpul moale, cu douǎ aripi la indivizii inripati, cele fixate la mezotorace; a doua pereche de aripi este reprezentatǎ printr-o pereche de balansiere sau haltere, care se gǎsesc la toate dipterele adulte, chiar cand lipsesc aripile anterioare.
Halterele au o pozitie lateralǎ pe cel de-al treilea segment al toracelui.Aparatul bucal este adaptat pentru supt sau pentrau intepat si supt.Tarsele sant alcǎtuite din cinci articole.
Corpul este alcǎtuit din cap, torace, si abdomen, cel mai adesea legate intre ele prin portiuni subtiri.
Capul este bine separat de torace si asezat pe un gat subtire, cu unele exceptii, ca la hipoboscide, la care capul este strans apropiat de torace.La cap prezintǎ doi ochi mari care ocupǎ o mare parte a capului.Acestia sant fatetati si adesea ei sant acoperiti cu o pubescentǎ finǎ.Pe vertex se gǎsesc trei oceli.Intre ochi sant inserate antenele care la diptere nematocere sant lungi si formate din multe articole, iar la dipterele brahicere sant foarte scurte si formate din trei articole principale, cel de-al treilea purtand un al patrulea articol, care seamanǎ cu un pǎr simplu; adesea acest al patrulea articol sǎ fie segmentat.
La partea inferioarǎ a capului se afla baza aparatului bucal, care are formǎ de trompǎ, mai mult sau mai putin alungitǎ.
Protoracele este de regulǎ putin dezvoltat si de asemenea si metatoracele, incat mezotoracele este rare cea mai mare a toracelui si cea mai vizibilǎ asemanator cu al himenopterelor. Segmentele toracelui, ca si abdomenul, pot sǎ fie acoperite de peri sau de solzisori cu o aranjare caracteristicǎ.
Toracele poartǎ trei perechi de picioare, tarsul avand intotdeauna cinci articole, primul fiind numit metatars, iar ultimul care poartǎ si ghearele , fiind numit si distars, praetars, sau onychium.Ultimul articol poartǎ douǎ gheare precum si niste formatii adezive, ca pernite sau peri (pulvili,arolium,empodium).
Aripile sant foarte subtiri si au o nervatie caracteristicǎ pentru diferite grupe.O nervatie tipicǎ cuprinde in general putine nervuri, care iau diferite aspecte in cadrul ordinului.Balansierele au forma unor mici apendici, ca niste firisoare terminate la capǎt cu cate o mǎciuca aproape sfericǎ.Ele sant organe senzoriale si au rol in coordonarea zborului.
Abdomenul este vizibil inelat, segmentele sant flexibile, iar pe partile laterale poartǎ stigmele respiratorii.Abdomenul se terminǎ cu douǎ apendice dorsale(cercii) si douǎ apendice ventrale (stilii).Ultima parte a abdomenului este hipopigiul, iar la femele ovopozitorul (oviscaptul).
Dezvoltarea este holometabolǎ.Numǎrul ouǎlor variazǎ de la cateva sute (la Musca, la Culex) si chiar panǎ la cateva mii (la Echinomya- o tahinida).Marea majoritate a dipterelor sant ovipare, dar sant si multe forme vivipare, mai ales printre tahinide.
Larvele de diptere sant de douǎ tipuri, unele acefale, altele eucefale, la acestea din urmǎ capul fiind bine dezvoltat. Larva se transformǎ in cursul metamorfozei intr-o pupǎ liberǎ mobilǎ, ca la culicide, sau o datǎ cu transformarea larvei in pupǎ, tegumentul larvei se umflǎ si se intareste, formand un invelis ca un butoias, in care este inchisǎ pupa.
Ordinul Lepidoptera
Numele ordinului derivǎ de la gr.lepis,lepidos=solz; pteron=aripa, popular sant numite fluturi.
Corpul lepidopterelor este acoperit fie cu solzi de diferite forme,mai alungiti sau mai lati, sau cu peri care pot fi la unii foarte desi si lungi, formand tufe pe suprafata corpului.Solzii si perii au aceeasi origine epidermicǎ , solzii fiind niste peri latiti.Solzii se iau pe degete cand fluturele este apucat cu mana si apar ca un praf fin.Lepidopterele sant insecte cu culori extrem de variate, frumoase, adesea stralucitoare, metalice, mai ales la aripi.
Capul fluturilor este mic in comparatie cu restul corpului si la multi fluturi o mare parte din cap este acoperitǎ de ochi, restul suprafetei capului fiind acoperitǎ cu peri.Intre ochi se distinge fruntea.In partea superioarǎ a fruntii sant inserate antenele.Antenele au forme variate, acestea utilizandu-se si pentru clasificatie.La multe forme, antenele sant filiforme, fie mai umflate la varf - claviforme, fie mai umflate la mijloc- fusiforme.
In general, antenele sant cilindrice sau putin turtite, dar la unele forme pot fi prismatice, cu sectiune triunghiularǎ.Sant antene care au la articolele lor cate un mic dinte lateral- antene dintate, altele au cate o micǎ prelungire ca niste lame sau ca niste dinti de pieptene- antene pectinate; sant si antene care au pe articolele lor fire lungi si subtiri, antene semǎnand cu o panǎ- antene plumoase.
Ochii sant fatetati, numǎrul fatetelor fiind foarte mare la unele forme (2700 la Sphinx).Ochii pot fi inconjurati cu cili lungi sau pot chiar sǎ poarte pe suprafata lor perisori.La unii fluturi se aflǎ cate un ocel in partea de deasupra a fiecarui ochi (la multi heteroceri).
La cap se distinge trompa, care in repaus stǎ strans in spiralǎ, intr-un sant care se aflǎ pe partea ventralǎ a capului.La unii fluturi trompa este foarte lungǎ, mai lungǎ chiar decat corpul (la unele sgingide), insa la altii trompa poate lipsi cu totul.
La fluturii primitive Micropterygidae exista un aparat bucal primitive , de tip ortopteroid, cu maxile alcstuite din cardo stipes, lacinia si galea, cu mandibule functionale.Palpii maxilari ai acestora sant alcatuiti din cinci articole iar labrum si labium sant reprezentate La fluturii evoluati, tipul de aparat bucal este asa numitul tip maxilar, la care cele mai multe piese s-au redus mult sau chiar au dispǎrut dezvoltandu-se numai maxilele.
La un astfel de aparat bucal distingem un labrum foarte mic, trompa si palpii labiali bine dezvoltati, apoi palpii maxilari sant foarte rudimentari, din 1-3 articole.Trompa este formatǎ din jgheaburi lungi care imbucandu-se cu marginile lor, formeazǎ tubul trompei.Aceste douǎ jgheaburi lungi reprezintǎ lobii externi ai maxilelor , mult alungiti - galea.
Toracele are o segmentatie foarte heteronomǎ, protoracele fiind intotdeauna mic, mai mult sau mai redus, iar mezo si metatoracele sudate, mezotoracele fiind intotdeauna mai mare.Picioarele sant in general homonome.
Coxele sant mari , iar trohanterele mici.Tibiile anterioare poartǎ adesea in jumǎtatea lor distalǎ un spin; tibiile mijlocii si cele posterioare poartǎ de regulǎ unul sau doi spini terminali si adesea si doi spini mediani sau superiori.
Tarsele sant alcǎtuite din cinci articole homonome,ultimul articol tarsal avand douǎ gheare si douǎ pulvile.Picioarele sant de regulǎ mici si subtiri si servesc mai mult insectei sǎ se prindǎ cǎnd se aseaza pe diferite corpuri si mult mai putin la mers.Aripile sant in numǎr de douǎ perechi. Ele sant de regulǎ bine dezvoltate , de cele mai multe ori aripile anterioare fiind mai mari decat cele posterioare, dar la multi fluturi ele sant aproape egale.
Coloratia aripilor este caracteristicǎ foarte evidenta a fluturilor, in mod obisnuit putandu-se recunoaste speciile dupǎ culoarea aripilor.
Schema nervatiei generale a aripilor aratǎ prezenta nervurilor longitudinale caracteristice si aripilor alte insecte: costalǎ, subcostalǎ, radiale, mediane, cubitale,anale.Medianele si cubitalele sant legate intre ele, in regiunea mijlocului aripii, prin nervure transversale , atat la aripa anterioarǎ cat si la cea posterioarǎ, formand astfel la fiecare aripǎ cate o celulǎ mare, numitǎ celula mediana sau discalǎ.La cei mai multi fluturi, nervatia aripilor posterioare diferǎ de aceea a aripilor anterioare, dar la altii este aceeasi nervatie.
Abdomenul este format din zece segmente, insa la mascul sant vizibile numai opt,deoarece sternitul segmentului 1 este rudimentar sau chiar lipseste, iar sternitele 9 si 10 formeazǎ la mascul aparatul copulator.Acest aparat este de regulǎ ascuns in gfsegmental 8.La femelǎ se vǎd sapte segmente, deoarece segmentele 8,9, si 10 sant retrase in segmentul 7 si nu se mai vǎd la exterior decat partial sau de loc.
La locul de unire dintre torace si abdomen se aflǎ la cele mai multe lepidoptere , o pereche de organe, situate de regulǎ pe torace, cunoscute sub numele de timpane, care se crede cǎ au rol auditiv, deoarece au fost descries organe cordotonale ce sant in legaturǎ cu aceste timpane.
Fluturii au metamorfoza completǎ- holometabolǎ.Larvele lor sant cunoscute sub numele de omizi.Forma si infǎtisarea omizilor este foarte variatǎ, totusi ele au caractere generale comune, si anume: corp in general cilindric,cu cinci panǎ la opt picioare.Cele mai multe au opt perechi de picioare si anume:trei perechi de picioare articulate, pe segmentele toracice si trei perechi de picioare simple, nearticulate, pe segmentele abdominale 3,4,5,6,10.Omizile au cinci perechi de picioare, care se mai numesc si geometride sau cotari, au trei perechi de picioare articulate, toracice, si douǎ perechi de picioare nearticulate, situate la capǎtul posterior al abdomenului.
Picioarele abdominale se mai numesc si picioare membranoase, picioare false sau pedesspurii.Aceste picioare abdominale poartǎ la capatul lor cate o coroanǎ de carlige, mai mult sau mai putin completǎ.
Pupa lepidopterelor este o pupǎ de tipul pupǎ obtectǎ.Ele au tecile aripilor, picioarelor si antenele vizibile, des strans lipite de corp,fiind invelite cu o membrane foarte finǎ. Sant insǎ si multe forme de lepidoptere cu aceste apendice libere, pupa acestora asemǎnandu-se cu cea de trichoptere.
Si unele lepidoptere superioare au pupa liberǎ, cum sant nepticulidele, heliozidele.Pupele au unele segmente ale corpului mobile; segmentele 8,9 si 10 sant totdeauna fixe, toate celelalte segmente putand fi mobile la unele forme si fixe la celelalte.
Fluturii sant unii diurni, altii nocturni sau crepusculari.Viata lor este scurtǎ, adesea de cateva zile , cel mult cateva sǎptamani.
Fluturii se hrǎnesc sugand diferite lichide, ca nectar din flori, seva ce se scurge din rǎnile plantelor, apa din sol, unii chiar lichide din gunoaie, din bǎligar si din dejectii, cum sant multe nimfalide.
La multi fluturi se observǎ dimorphism sexual, fie in deosebirea de mǎrime intre mascul si femela, fie dupǎ culoare, dupǎ aspectul general al corpului, dupǎ antene.
Marea majoritate a fluturilor au o singurǎ generatie pe an, dar sant unii care se dezvoltǎ foarte repede, avand 2-3 sau chiar mai multe generatii intr-un an.Sant si forme a cǎror dezvoltare completǎ se face in mai multi ani.
Ordinul Dermaptera
Insecte cu infǎtisare caracteristicǎ, usor de deosebit de alte grupe de insecte prin corpul lor alungit, turtit dorso-ventral, totdeauna de culoare brunǎ-castanie, cu aripi scurte care lasǎ mai multe segmente abdominale descoperite.
Capul este turit dorso-ventral.Poartǎ doi ochi mici, lateral spre partea anterioarǎ.Nu au oceli.Antenele sant lungi si filiforme.Toracele este alcǎtuit dintr-un protorace mobil, care este de mǎrimea capului, turtit dorso-ventral si putandu-se misca liber fata de mezotorace si de mezo si metatorace acoperite de elitre pieloase si scurte.
Aripile membranoase sant ascunse in intregime dedesuptul primei perechi de aripi; aceste aripi membronoase nu sant totusi aripi mici si reduse, ci dimpotrivǎ, cand se intind sant mari, cu nervuriler asezate radiar, ca la un evantai.Ele se strang sub aripile pergamentoase, ca un evantai ,in jurul unui punct de la marginea anterioarǎ, apoi indoindu-se inca de douǎ ori transversal.Cu toate aceste aripi membranoase dezvoltate, foarte putine sant formele care zboarǎ, de regulǎ ele sant insecte alergǎtoare, fugind foarte repede.Sant si forme care au aripile rudimentare sau le lipsesc total.
Piesele bucale sant de tip ortopteroid si sant masticatoare.Abdomenul este foarte mobil, vizibil inelat, cu tegumentul lucios.Abdomenul este alungit si format din 9 segmente vizibile la care se adaugǎ si segmentul anal.La femelǎ, segmentele 7 si 8 sant foarte reduse si nu se vǎd decat 7 segmente.La extremitatea abdomenului se aflǎ o pereche de cerci mari sub forma a doua apendice uniarticulate, mai dezvoltate la mascul decat la femela.La mascul,aceste pense sau forcepsuri sunt mai curbate si poartǎ dinti la baza, pe parte internǎ; la femela forcepsurile sant drepte si necurbate.Totusi aceste cǎngi variazǎ dupǎ formele considerate,ceea ce poate servi drept caractere specifice.Aceste cǎngi au mare asemǎnare cu cele de la Japygidae.Se considerǎ ca aceste forcepsuri ar avea rol de apǎrare.
Forficulidele sant insecte nocturne, ziua stau ascunse prin crǎpǎturile pǎmantului, pe sub bulgǎri, pe sub pietre, sub trunchiuri, sub scoarta, in frunzar.
Regimul lor alimentar este totusi carnivor,mancand larve, melci; uneori atacǎ si fructele cǎzute din arbori.
Primavara depun ouǎ in pǎmant, aproape de suprafatǎ, intr-o mica excavatie; femela aparǎ aceste ouǎ panǎ cand apar larvele.Metamorfoza este paurometabola, larvele semǎnand mult cu adultii, dar fiind lipsite de aripi.
Ordinul HETEROPTERA
Insectele cuprinse in acest ordin au patru aripi; prima pereche de aripi este intǎritǎ la partea lor bazalǎ si membranoasǎ la varf, iar perechea a doua este membranoasǎ in intregime.Aparatul bucal este conformat pentru intepat si supt, baza rostrului aflandu-se in parte anterioarǎ a capului, departe de coxele primei perechi de picioare.Se dezvoltǎ prin metamorfozǎ gradatǎ.
Insecte foarte comune, mult rǎspandite ,se intalnesc pe florile plantelor,pe frunze, pe fructe;unele trǎiesc pe suprafata apelor sau in apǎ.Putine forme duc o viatǎ parazitarǎ pe pǎsǎri sau pe mamifere.Putine heteroptere pot fi considerate insecte utile,dupǎ obiceiurile lor de animale prǎdǎtoare care atacǎ unele insecte dǎunatoare.
Numele lor provine de la structura aripilor anterioare care au partea bazalǎ mai intǎritǎ, cornoasǎ numitǎ hemielitrǎ.
Aceste aripi nu servesc la zbor, ci au un rol numai de a proteja aripile posterioare,membranoase, care sant stranse in lung dedesuptul hemielitrelor.Partea bazalǎ a aripilor anterioare se numeste corium, iar partea terminalǎ membranǎ.Cele douǎ hemielitre sant separate la baza lor prin scutel, care are formǎ triunghiularǎ, totdeauna dezvoltat, uneori chiar prelungindu-se lateral ca o lamǎ care poate acoperii complet elitrele.
Structura hemielitrei mai cuprinde si alte particularitati caracteristice:la partea internǎ prezintǎ o lamǎ cu marginile paralele legate de corium printr-o suturǎ flexibilǎ care se numeste clavus.Unele familii prezintǎ la partea anterioarǎ a coriului, langǎ membrane, o piesǎ mica triunghiularǎ, cuprinsǎ chiar in corium, subtire si mobile , numitǎ cuneus.La multe heteroptere, corium prezintǎ o lama cornoasǎ si in lungul marginii anterioare a hemielitrei numitǎ embolium.
Partea de la varf a hemielitrei, membrane este subtire si prezintǎ nervuri si uneori si una sau mai multe celule.Dispozitǎia acestor nervuri si celule serveste in multe cazuri la determinarea formelor.Nervatia aripilor la heteroptere reprezintǎ o distanta insemnatǎ de tipul nervatiei primitive.Pot exista chiar la aceeasi specie indivizi cu hemielitrele si aripile bine dezvoltate (forme macroptere) si altele la care hemielitrele sant foarte reduse (forme brahiptere).Unele forme au membrane hemielitrelor foarte redusǎ sau chiar complet redusǎ si de asemenea si aripile posterioare.
Antenele au 4-5 articole,rostrul in general alcǎtuit din 4 articole mai rar din 3.Dupǎ cap urmeazǎ corseletul sau pronotul,uneori cu formǎ patrulaterǎ ,alteori trapezoidal uneori avand pe laturi un dinte, un spin, un unghi.
Abdomenul prezintǎ pe partea ventrala 6 segmente bine vizibile.Marginile laterale ale segmentelor abdominale sau in general foarte subtiate si cu forma unei lame marginale.Multe heteroptere rǎspandesac un miros neplǎcut produs de o glandǎ mirositoare, care se deschide pe metatorace, aproape de baza picioarelor posterioare.
Ordinul Trichoptera
Insecte aripate, ale cǎror aripi sunt membranoase, totdeauna mai mult sau mai putin cu peri lungi mǎtǎsosi. La cele mai multe forme nervatia este de tip primitiv, dar cu putine nervuri accesorii. La unele forme specializate nervatia aripilor este redusǎ Piesele bucale sant mult reduse, palpii fiind mai dezvoltati. Dezvoltarea este holometabolǎ.
Sant de regulǎ mai mult cunoscute dupǎ larvele lor, care se intalnesc adesea in mare numǎr in bǎlti si rauri si se recunosc dupa tuburile lor caniste mansoane fǎcute din diferite materiale din apǎ, ca nisip, bucati de plante, resturi de scoici. Adultii sunt de regulǎ nocturni, ziua stand pe plante, pe frunze, si ramuri. Au culori sterse, de regulǎ cu nuante brune si galbene.
Aparatul bucal este de tip masticator, dar deseori este rudimentar. Palpii maxilari, ca si cei labiali sant bine dezvoltati. De regulǎ palpii maxilari au 5 articole.
Picioarele au o conformatie normalǎ , iar tarsele sant alcǎtuite din 5 articole. Tibiile si tarsele poartǎ spini caracteristici.
Ambele perechi de aripi sant membranoase si acoperite de o pubescentǎ deasǎ, finǎ si mǎtǎsoasǎ , culoarea acestor peri dand si coloratia aripii, petele si desenele de pe aripi, mai ales cele de pe partea anterioarǎ. Nervatia aripii este foarte caracteristicǎ si prezintǎ particularitati la diferitele specii, servind astfel la determinrea formelor. Determinarile se fac cu rigurozitate dupǎ armatura genitalǎ a masculilor, care au insa piese foarte fine si foarte fragile si destul de greu de observat .
Adultii de regulǎ zboarǎ ziua, majoritatea zboarǎ seara si noaptea, zborul lor fiind foarte incet si destul de scurt. Ei trǎiesc putin, de regulǎ numai cateva zile si foarte rar 2-3 sǎptamani. Ei nu se hrǎnesc in acest timp si numai unoeri ling nectar din flori sau picǎturi de apǎ de pe frunzle plantelor.
Larvele sunt de douǎ tipuri : unele sant de tip campodeiform si se intalnesc la familiile Rhyacophilidae si Hydropsychidae. Ele nu isi fac tub, ci se misca libere in apǎ. Unele dintre acestea tes in apǎ un fel de plase din fir de mǎtase cu ajutorul cǎrora prind mici animale acvatice. Alte larve sant de tip eruciform, cu capul de regulǎ ortogonat.
Ele au apendice abdominale pe primul segment abdominal, nearticulate, iar pe ultimul segment abdominal o alta pereche de apendice, articulate si purtand niste gheare si carlige de diferite forme. Cu aceste apendice abdminale de pe primul si ultimul segment abdominal larvele se tin bine agǎtate, in interiorul tubului. De fapt si larvele fǎrǎ tuburi au apendice abdominale , dar ele sant reduse.
Toate larvele de trichoptere sant acvatice, o singurǎ formǎ este terestrǎ , Enoicicla pusilla Burm., care isi face un tub din nisip si resturi de plante si traieste tot prin apropierea apelor, prin muschi.
Biologia larvelor de trichoptere , ca si a nimfelor lor, este deosebit de interesantǎ mai ales prin marea variatie a tuburilor larvare in privinta formelor lor si a materialelor din care sant construite . De regulǎ, fiecare specie are un anumit model de tub, dar destul de des infatisarea tubului, dar mai ales materialele dincare el este construit , variazǎ la aceeasi specie dupa materialele pe care larva respectivǎ le aseste in apǎ in care traieste si se dezvoltǎ. De asemenea, infatisarea tubului poate sǎ difere si cu varsta larvei.
Cand larva a ajuns in stadiul matur, ea se pregateste sǎ se transforme in nimfǎ. Larvele care trǎiesc in tub isi fixeaza tubul pe un corp din apa lipindu-l cu cu fire de mǎtase. De regulǎ larva rupe capǎtul posterior al tubului, scurtǎnu-l si facandu-l sǎ mai aibǎ astfel inca o deschidere , pe langa deschiderea anterioarǎ. Larvele inchid apoi cele douǎ deschideri , lǎsand numai crǎpǎturi pe unde sǎ intre apa.
In felul acesta tubul larvar devine un adapost nimfal. Celelalte larve libere , fǎrǎ tub , isi construiesc si ele o casuta de nisip si pietrecele, pe care lipesc de o piatrǎ mai mare din apa .In inchisorile lor, larvele isi tes o captuseala de mǎtase. Larvele de trichoptere servesc in mare masura la hrana pestilor.
Biologia nimfelor are drept caracteristic faptul ca ele rǎman tot acvatice, ceea ce nu se intamplǎ cu alte larve acvatice, in afara de unele diptere; in al doilea rand, aeste nimfe nu sant imobile, ci ele se miscǎ si se agitǎ in casuta lor. Nimfa lor este de tip pupǎ liberǎ.
Nimfa are o duratǎ a stadiului nimfal de douǎ sǎptamani. Cand nimfa s-a apropiat de terminarea stadiului nimfal, ea iese din adǎpostul sau se duce din nou o scurtǎ perioada de viatǎ liberǎ, dupǎ care nǎparleste si din ea iese adultul. Nimfa iese din casutǎ, ea rupe invelisul care o tinea inchisa, cu ajutorul mandibulelor sale, care sant puternice. Nimfa devenitǎ liberǎ inoatǎiin apa cu picioarele, se tareste pe un suport oarecare si iese din apǎ; aici tegumentul nimfei se usucǎ, se produce o crǎpǎturǎ pe cap si pe torace si adultul iese acu mdin exuvia nimfei. La unele forme de trichoptere nimfa nu se tarǎste afarǎ din apǎ pe un suport oarecare, ci inoatǎ la suprafata apei, scotand din apa partea anterioarǎ a corpului; tegumentul se usucǎ acolo si iese imago.
Clasificatia tricopterelor a suferit numeroase modificari, ca multe alte clasificatii entomologice. O clasificatie admisǎ mai recent este in doua subordine cu un total de 13 familii: Subordinul Inaequipalpia si Subordinul Aequipalpia.
Ordinul Orthoptera
Acest grup de insecte se caracterizeazǎ prin insecte de mǎrimi mijlocii sau mari. Ele au de regulǎ capul ortogonat, mai rar hipogonat. Au doi ochi fatetati si cel mai adesea si trei oceli . Aparatul bucal este de tip primitiv masticator.
Antenele sant de tip setacee sau filiforme, mai rar mǎciucate. Unele forme au chiar antene turtite.
Prototoracele este bine dezvoltat, cu pronot neted sau cu carene.Au douǎ perechi de aripi, prima pereche fiind pergamentoase si sant numite elitre sau tegmine; ele sant mai inguste decst cele posterioare si au nervuri.Aripile posterioare sant late si sant stranse in falduri longitudinale in lungul corpului. Multe forme au aripi rudimentare.
Picioarele sant heteronome, cele posterioare fiind adaptate la sǎrit, iar unele forme avand picioarele anterioare adaptate la sǎpat. Tarsele au 3-4 articole , mai rar 5. Organele stridulante se aflǎ la baza aripilor sau pe femurele posterioare.
Organele timpanale sant situate pe tibiile anterioare sau la baza abdomenului, la unele forme lipsesc total .
Abdomenul prezintǎ zece segmente si o pereche de cerci la ambele sexe. Femelele prezintǎ totdeauna oviscapt. Acesta este alcǎtuit din trei perechi de teci lungi, formand un oviscapt lung, cu forma de sabie, sau din trei perechi de teci scurte si indoite la varf ca niste cǎngi (la Acrididae), fǎrǎ sǎ formeze un oviscapt lung ca la Tettigoniidae. Oviscaptul la Gryllidae este format din douǎ perechi de teci, care impreunǎ formeazǎ un fel de tub lung cilindric.

Lista taxonomica a speciilor de lepidoptere nocturne din probe
Familia Cossidae
Phragmatecia castanea Hb.
Familia Hepialidae
Hepialus sylvinus L.
Pterophoridae
Pterophorus pentadactyla L.
Capperia britanniodactyla
Alucita hexadactyla
Gracillariidae
Cameraria ohridella D. et D.
Phylonorycter robiniella Clemens
Pyralidae
Hypsopygia costalis Fabr.
Pyrausta aurata Scop.
Pyrausta purpuralis L.
Pleurota ruralis Scop.
Paraponyx stagnata D.
Onocera semirubella Scop.
Eurrhypara hortulata L.
Crambus pratella L.
Pyrausta aurata Scop.
Ostrinia nubilalis Hb.
Lymantriidae
Lymantria disparL. (f. melanica)
Arctornis l-nigrum Mull.
Arctiidae
Miltochrista miniata Forst.
Oenistis quadra L.
Phragmatobia fuliginosa L.
Lithosia lutarella L.
Panaxia quadripunctaria Poda
Drepanidae
Cilix glaucata scop.
Drepana harpagula Esp.
Drepana binaria Hufn.
Axiidae
Habrosyne pyritoides Hfn.
Notodontidae
Pterostoma palpina Cl.
Habrosyne derasa L.
Clostera anastomosis L.
Odonestis pruni L.
Pygaera anastomosis L.
Spatalia argentina D. et S.
Syntomidae
Dysauxes ancilla L.
Noctuidae
Abrostola triplasia L.
Aedia leucomelas L.
Amphipyra livida D. et S.
Amphypyra pyramidea L.
Apamea monoglypha Hfn.
Asteroscopus sphinx Hfn.
Agrochola circellaris Hfn.
Conistra veronicae Hb.
Autographa confusa Hb.
Autographa gamma L.
Calophaena opalina
Catocala elocata Esp.
Catocala nupta L.
Colobochyla salicalis D. et S.
Cosmia pyralina D. et S.
Cucullia fraudatrix Ev.
Deltote bankiana Fabr.
Diachrysia chrysitis L.
Diachrysia chryson Esp.
Elaphria venustula Hb.
Erastria trabealis Scop.
Eucarta virgo Treit.
Gonospileia glyphica L.
Helicoverpa armigera Hb.
Helicoverpa peltigera D. et S.
Heliothis cardui Hb. (V. Ghiunaltu)
Heliothis maritima bulgarica Draut.
Hylophila prasinana
Hypena proboscidalis l.
Lygephila craccae D. et. S.
Melanchra persicariae L.
Mycteroplus puniceago Bsdv.
Mythimna congrua Hb.
Naena typica
Noctua fimbriata Schreber
Noctua pronuba L.
Prodotis stolida Fabr.
Protoschinia scutosa D. et S.
Pseudeustrotia candidula D. et S.
Pseudoips fagana fabr.
Pyrrhia umbra Hfn.
Rusina ferruginea Esp.
Scoliopterix libatrix L.
Sideritis conigera D. et S.
Tyta luctosa D. et S.
Toxocampa craccae F. (?)
Trachea atriplicis L.
Ulochlaena hirta Hb.
Viminia rumicis L.
Xestia c-nigrum L.
Xylina vetsusta L. (Nifon)
Geometridae
Operoptera brumata L.
Anaitis plagiata L.
Anticlea derivata D. et S.
Aspilates gilvaria D. et S.
Boarmia rhomboidaria D. et S.
Boarmia selenaria Schiff.
Calothysanis amata L.
Campaea margaritata L.
Camptograma bilineata
Chlorissa viridata L.
Cidaria ocellata L.
Cosmorhoe ocellata L.
Cosymbia orbicularis
Cosymbia punctaria L.
Cosymbia pupillaria Hb.
Cyclophora albipunctata Hb.
Cyclophora annulata Sch.
Ematurga atomaria L.
Endrosis sarcetella (de verificat) - 27.09.
Ennomos erosaria D. et S.
Ennomos quercinaria Hfn.
Epione repandaria Hfn.
Euchloris smaragdaria fabr.
Euphitecia centaureata D. et S.
Euphitecia extraversaria H.S.?
Euphitecia linariata F.
Euphitecia pyreneata Mab.
Hemistola chrysoprasaria Esp.
Hipparchus papilionaria L.
Horisme corticata Treit.
Horisme terasata
Idaea muricata Hfn.
Ligidia adustata D. et S.
Lomaspilis marginata L.
Lythria purpuraria L.
Mesoleuca albicillata L.
Perizoma lugdunaria H.S.
Plagodis dolabraria L.
Rhodostrophia calabra Petagna
Scopula immorata L.
Scopula rubiginata Hfn. (?)
Selenia tetralunaria Hfn.
Semiothisa clathrata L.
Semiothisa notata L.
Sterrha aversata L.
Sterrha ochrata Scop.
Tephrina arenacearia Hb.
Thalera fimbrialis Scop.
Sphingidae
Macroglossum stellatarum L.
Laothoe populi L.
Marumba quercus L.
Deilephila elpenor L.
Celerio galii Rott. (larva)
Celerio euphorbiae L.(larva)
Fam. Saturnidae
Saturnia pyri L.
Prezentarea principalelor specii de lepidoptere nocturne; Fam. Geometridae, Noctuidae, Sphingidae
1.1Fam.Geometridae
Genul Chlorissa
Chlorissa viridata Linnaeus 1758
Perioada de zbor: jumǎtatea lunii mai si jumatatea lunii iulie.
Biotop: locuri deschise cu multe buruieni, luminisuri, liziere insorite.
Plante gazda: luminisuri cu vegetatie spontanǎ bogatǎ unde sunt prezenti arbori ca: Betula sp, Quercussp, Corylus avellana, Salix sp, Rubus fruticocus.
Ponta si dezvoltarea: in decada a doua a lui mai femelele depun ouǎle pe plantele gazdǎ. Larva si crisalida se hraneste activ dupǎ eclozare panǎ in august cand isi face un cocon in care se impupeaza, iernand in acest stadiu. In mai eclozeaza adultul .
Raspandire: prezentǎ in Europa si in Romania.
Genul Hemistola
Hemistola chysoprasaria Esper 1794
Perioada de zbor:sfarsitul lunii iunie panǎ in prima jumatate a lunii august
Biotop: margini de pǎduri, zone cu tufisuri, grǎdini, parcuri, in general preferǎ locurile calde.
Plante gazda: Clematis vitalba, C. viticella
Ponta si dezvoltarea: femelele depun ouǎle de obicei in luna iulie astfel ca omizile se pot observa din august; spre sfarsitul toamnei intra in diapauza din care ies in urmatorul an; se impupeazǎ si spre sfarsitul lui iunie eclozeaza adultii.
Rǎspandire: prezentǎ in Europa si in Romania.
Genul Cyclophora
Cyclophora annulata, Schulze 1775
Perioada de zbor: prezintǎ douǎ generatii: prima generatie incepe la sfarsitul lunii aprilie panǎ in prima jumatate a lunii iunie: a doua generatie tine de la sfarsitul lunii iunie panǎ la sfarsitul lunii august.
Biotop: in pǎduri de foioase
Plante gazda: Fagus sylvatica, Acer campestre. Betula pendula,Carpinus betulus.
Ponta si dezvoltarea:
ouǎle primei generatii in luna mai , iar in generatia a doua in
luna august.Larva eclozeazǎ in iunie, se impupeaza in iulie intre
Rǎspandire: prezentǎ mai ales in Europa Centrala si de Sud, de asemenea si in Romania.
Genul Scopula
Scopula immorata, Linnaeus 1758
Perioada de zbor: sfarsitul lunii mai panǎ la sfarsitul lunii iulie
Biotop: ierburile din locurile necultivate, locuri aride, marginea livezilor, a pǎdurilor, turbǎrii, luncii, zone innierbate.
Plante gazdǎ: Artemisia vulgaris, Achillea millefolium, Thymus serpyllum, Calluna vulgaris, Salvia pratensis.
Ponta si dezvoltarea: din crisalida ce a iernat eclozeaza fluturii. Dupǎ imperechere femelele depune ouǎle pe plantele preferate incepand din luna mai panǎ in luna iulie, dupǎ mai multe zile in luna august eclozeazǎ larvele ce sunt active panǎ toamna tarziu cand intra ǎn diapauza hibernalǎ.
Rǎspandire: prezentǎ in Europa si in Romania
Scopula rubginata , Schrank 1802
Perioada de zbor: prezintǎ douǎ generatii: prima jumatate tine de la prima jumǎtate a lunii mai inceputul lunii iulie; iar a doua generatie tine de la prima jumǎtate a lunii iulie panǎ la prima jumatate a lunii septembrie.
Biotop: locuri necultivate cu florǎ spontanǎ , coaste insorite, bǎlǎrii etc.
Plante gazdǎ: Trifolium sp, Lotus sp, Vicia sp, Polygonium aviculare, Medicago sp, si diverse plante mai ales leguminoase.
Ponta si dezvoltarea: in Europa de Nord depune ponta in luna iulie,eclozeazǎ larva dupa 7-8 zile , ierneaza si-isi defineste dezvoltarea in primavara urmǎtoare impupandu-se in luna mai si iunie, iar eclozare adultului este in iunie. Larva deranjatǎ se strange in jurul sǎu formand douǎ bucle caracteristice. Larva se impupeaza intr-un cocon la suprafata solului sǎu sub muschi. In cazul in care are mai multe ganeratii pe an larval se pot observa din luna aprilie- mai panǎ in luna septembrie.
Raspandire: prezentǎ in Europa in afarǎ de zonele nordice, de asemenea prezentǎ si in Romania.
Genul Idaea
Idaea muricata, Hufnagel 1767
Perioada de zbor: inceputul lunii iunie - sfarsitul lunii august.
Biotop: pajisti umede, liziere de pǎduri cu ierburi, locuri sǎrǎcǎcioase, uneori in locuri aride, nisipoase cu plante.
Plante gazda: Platago lanceolata, Polygonium sp, Galium palustre, Calluna vulgaris.
Ponta si dezvoltarea: femelele depune ouǎle intr-un interval mai mare de timp.Dupǎ eclozare larva se hraneste panǎ in luna iulie apoi intrǎ in diapauaza hibernalǎ , cautandu-si dezvoltarea, in primavara urmǎtoare, impupandu-se spre sfarsitul lunii mai, iar in iunie eclozeaza adultul.
Rǎspandire: prezentǎ in Europa si in Romania, in zone preferate.
Genul Lythria
Lythria purpuraria, Linnaeus 1761
Perioada de zbor: prezintǎ douǎ generatii: prima generatie incepe in prima jumǎtate a lunii aprilie panǎ la sfarsitul lunii iunie; a doua generatie incepe in luna iunie panǎ la inceputul lunii septembrie
Biotop: pante si vǎi pietroase, calde , pasuni, pǎmanturi intelenite, miristi si alte locuri calde.
Plante gazda: Polygonium aviculare, preferand flori si fructele.
Ponta si dezvoltarea: femelele depune ouǎle in luna mai -iunie si iulie august, ierneaza ca pupǎ. In regiunile nordice poate avea numai o generatie.
Rǎspandire: prezentǎ in Europa si in Romania; mai multe forme in functie de generatie si de zona climaticǎ.
Genul Camptograma
Camptogramma bilineata bilineata
Perioada de zbor: incepand din luna mai pana la inceputul lunii august.
Biotop: grǎdini, parcuri, turbǎrii, margini de pǎdure, in zone cultivate, locuri virane practice aproape peste tot.
Planta gazda: Stellaria media, Rumex sp, Taraxacum sp, Fragaria vesca, Potentila sp, Urtica sp, Plantago sp., si de alte specii de plante joase
Ponta si dezvotarea: femelele depune sub formǎ neregulatǎ
ouǎle pe
Rǎspandire: toatǎ Europa si in Romania.. Avand o rǎspandire largǎ are mule forme mai ales de culoare.
Genul Cosmorhoe
Cosmorhoe acellata, Linnaeus 1758
Perioada de zbor: prezintǎ douǎ generatii: prima generatie incepe in luna mai si tine panǎ la jumǎtatea lunii iunie; iar a doua generatie incepe la jumǎtatea lunii iulie si tine panǎ in luna septembrie.
Biotop: margini de pǎduri, turbǎrii, campie, grǎdini in functie de planta gazdǎ.
Planta gazda: Gallium sp.
Ponta si dezvoltarea: femelele depune ouale in luna mai si in luna iulie generatia a doua, larvele observabile din luna iunie-septembrie, generatiile adesea suprapunandu-se. Ierneaza ca larvǎ insa si ca crisalidǎ, in functie de conditiile anului respective.
Rǎspandire: in toata Europa si in Romania in functie de rǎspandirea plantei gazda.
Genul Horisme
Horisme tersata tersata, Denis &Schiffermuller 1775
Perioada de zbor: prezintǎ douǎ generatii: prima generatie incepe la sfarsitul lunii aprilie si dureazǎ panǎ la sfarsitul lunii iunie; iar a doua generatie incepe in luna iulie si tine panǎ la sfarsitul lunii august.
Biotop: margini de pǎduri, ǎi pante cu tufisuri intotdeauna pe terenuri calcaroase
Plante gazda: Clematis vitalba, C. viticella, Anemone sylvestris, Ranunculus acris.
Ponta si dezvoltarea: in locurile unde sre doua generatii femelele depune ouǎle In luna mai si iunie, larvele se dezvoltǎ in iunie -iulie, eclozand fluturii la inceputul lui iulie, astfel ca femelele depune ponta in luna august si larvele de dezvolta panǎ in septembrie cand se impupeazǎ. Crisalida este netedǎ de culoare brun-roscatǎ intr-un cocon in sol. Adesea generatia a doua este incompletǎ.
Rǎspandire: aproape in toatǎ Europa, exceptand regiunile nordice si de asemenea prezentǎ in Romania.
Genul Perizoma
Perizoma lungdunaria
Perioada de zbor: prima jumǎtate a lunii iunie panǎ in luna august.
Biotop: locuri insorite din zǎvoaie, tufisuri, buruieni, maluri de rauri si fluvii cu vegetatie.
Planta gazda: Cucublus baccifer si fructe sub forma de baca.
Ponta si dezvoltarea: femelele dupǎ eclozare in perioada de zbor depune ponta pe plantele preferate de larve ca hranǎ; in septembrie se impupeaza si ierneaza in stadium de crisalidǎ.
Rǎspandire: prezentǎ in Europa si in Romania, specie mai rarǎ.
Genul Eupithecia
Eupithecia linariata, Denis &Schiffermuller 1775
Perioada de zbor: prezintǎ douǎ generatii: prima generatie incepe la sfarsitul lunii mai panǎ la prima jumǎtate a lunii iulie: iar a doua generatie incepe la sfarsitul lunii iulie panǎ la inceputul lunii septembrie.
Biotop: zona de campie, terenuri intelenite, margini de drumuri mai putin circulate.
Plante gazda:Digitalis purpurea, Linaria vulgaris si Linaria sp se hrǎnesc cu flori si seminte.
Ponta si dezvoltarea:in cazul in care sunt douǎ generatii pe an femelele spre sfarsitul lui iulie uneori august iar cu o singurǎ generatie pe an femelele depune ouǎle in luna iulie, larva se dezvoltǎ in luna august-septembrie, impupandu-se la sfarsitul lui septembrie, , adultul eclozeazǎ in luna iunie anul viitor.Coconul alcaǎtuit din pǎmant, crisalida avand culoare brun-deschis.
Rǎspandire: aproape in toata Europa si Romania.
Eupithecia centaureata, Denis& Schiffermuller 1775
Perioada de zbor: prezintǎ douǎ generatii: prima generatie incepe in luna aprilie-mai si dureazǎ panǎ la prima jumǎtate a lunii iulie; iar a doua generatie incepe din luna iulie si tine panǎ in prima jumǎtate a lunii septembrie.
Biotop: margini de pǎduri, pǎdurile rare cu poieni, zone cu tufisuri.
Plante gazda: Achillea millefolium, Eupatorium cannabinum, Centaurea nigra, Senecio jacobaea, Solidago virgaurea, de asemenea si alte specii din genurile:Centaurea sp., Senecio sp., Solidago, Pimpinella sp., Peucedanum sp. s.a.Larva este polifagǎ preferand florile plantelor joase.
Ponta si dezvoltarea: femelele din prima generatie depun ouǎle pe plantele preferate ca hrana de omizi incepand din mai iar femelele din generatia a doua din iulie.Ponta fiind depusǎ esalonat si eclozarea adultilor este la fel. Crisalida este brunǎ sau rosiaticǎ, cea a generatiei a doua ierneaza.
Rǎspandire: aproape in toatǎ Europa si in Romania.
Genul Semiothisa
Semiothisa notata, Linnaeus 1758
Perioada de zbor: prezintǎ douǎ generatii: prima generatie incepe din luna aprilie/mai si dureazǎ panǎ la jumǎtatea lunii iulie; iar a doua generatie tine de la jumǎtatea lunii iulie panǎ in luna septembrie.
Biotop: pǎduri cu subarboret bogat, poieni in paduri de foioase, mestecǎnisuri si turbǎrii, , margini de pǎduri, lunci, grǎdini.
Plante gazda: Betula pendula, Salix caprea, S. cinerea, Allnus glutinosa, Quercus sp.
Ponta si dezvotarea: femela depune ouǎle in luna iunie;omizile au o dezvoltare rapidǎ astfel ca in luna iulie- august eclozeazǎ fluturii. Femelele depune ouǎle in luna august si in septembrie si se pot observa omizile care la maturitate coboarǎ si se impupeaza in sol.Crisalida ierneazǎ, fluturii din prima generatie la sfarsitul lui aprilie inceputul lunii mai. In zonele mai nordice sau in cazul unor ani mai reci are numai o generatie pe an, cand adultul eclozeazǎ in luna mai/iunie.
Rǎspandire: mai ales in Centrul si Nordul Europei si in Romania.
Genul Tephrina
Tephrina arenacearia, Lempke 1937
Perioada de zbor: prezintǎ douǎ generatii: prima generatie tine din luna iunie panǎ in luna iulie iar a doua generatie incepe in luna iulie si dureazǎ panǎ la inceputul lunii septembrie
Biotop: raristi, tǎieturi si margini de pǎduri, tufǎrisuri, pajisti, stancǎrii inierbate etc.
Plante gazda: Coronilla varia
Ponta si dezvoltarea: se impupeaza in sol. Biologie insuficient cunoscutǎ.
Raspandire: specie comunǎ in Europa si in Roamania, preferand zonele sudice.
Genul Plagodis
Plagodis dolabraria, Linnaeus 1767
Perioada de zbor:incepe la sfarsitul lunii aprilie si tine panǎ in luna iulie
Biotop: Pǎduri si margini de pǎduri bine insorite, livezi, grǎdini
Plante gazda: Betula pendula, Quercus sp., Salix sp., Tilia sp., Fagus sylvatica, Prunus domestica.
Ponta si dezvoltarea: adultul eclozeazǎ in luna aprilie, iar femelele depune ouǎle in luna mai-iunie; omizile se hrǎnesc panǎ in luna iulie, coboarǎ de pe plantele gazdǎ pe sol unde sub muschi se impupeaza. Larvele in timpul repausului stau nemiscate, greu de observat, avand aspectul unei ramuri tinere.
Raspandire: comunǎ in Europa si in Romania.
Fam Sphingidae
Genul Marumba
Marumba quercus, Denis &Schiffermuller 1775
Perioada de zbor: din luna mai panǎ in luna iulie
Biotop: Pǎduri de stejar
Planta gazda: Quercus sp., insa prefera mai ales Q. ilex, Q. pedunculata.
Ponta si dezvoltarea: prezentǎ in zona de campie panǎ la 1500 m. Adultii zboarǎ numai noaptea.Preferǎ zone uscate cu stejari, biologie putin cunoscutǎ.
Rǎspandire:aproape in toatǎ Europa si in Romania, fiind o specie mai xerofilǎ.
Genul Macroglossum
Macroglossum stellatarum, Linnaeus 1758
Perioada de zbor: prezintǎ douǎ generatii: prima generatie incepe in luna mai si tine panǎ in luna iulie; iar a doua generatie incepe la sfarsitul lunii august si tine panǎ in luna octombrie.
Biotop: pante si locuri insorite, de asemenea margini de pǎduri si locuri pietroase, grǎdini, campuri necultivate cu vegetatie spontanǎ bogatǎ, zbor diurn viziteazǎ florile din grǎdini si cele spontane.
Plante gazda: Galium molugo, G.verum, Rubia pregrina, Stellaria sp.
Ponta si dezvoltarea: femelele depun ouǎle izolate pe muguri sau florile plantelor pe care le cerceteazǎ in lunile mai, iulie si august dupǎ 7-8 zile eclozeazǎ omizile, care se pot observa in mai-iunie si august-septembrie. Impuparea pe sol intr-un cocon subtire.Perioada pupalǎ dureazǎ cca 30 de zile. In cazul a douǎ generatii pe an adultii sunt prezenti din aprilie panǎ in septembrie.Biologie inca confuzǎ, putin cunoscutǎ.
Rǎspandire:specie comunǎ mai ales in Sudul Europei insa prezentǎ si in zone mai nordice datoritǎ exemplarelor migratoare, prezentǎ si in Romania. Se presupune ca limita nordicǎ a arealului ar fi Marea Nordicǎ.
Genul Deilephila
Deilephila elephenor elephenor, Linnaeus 1758
Perioada de zbor: incepe din prima jumǎtate a lunii mai si tine panǎ in luna iulie.
Biotop: grǎdini, campuri, pǎduri de luncǎ rare, margini de ape si plant cu flori mirositoare si bogate in nectar pentru hranǎ.
Planta gazda: Epilobium angustifolium, E.hirsutum, Galium sp, Lythrum salicaria, Oenothera sp, polygonium sp.
Ponta si dezvoltarea: femelele depune ouǎle in luna iunie, isolate sau in grupe mici, pe partea inferioarǎ a frunzelor. Omizile eclozeaza dupǎ cca douǎ sǎptamani. Impuparea in sol la micǎ adancime intr-o loja nimfalǎ facutǎ de larvǎ, la sfarsitul verii sau toamna ;eclozarea fluturilor in urmǎtorul an in luna mai/iunie. In cazul a douǎ generatii pe an din crisalidǎ eclozeazǎ fluturii in luna octombrie, insǎ numai o parte din ei. Biologia este necesarǎ sǎ fie aprofundatǎ. Larva posedand homocromie este greu observabilǎ, insǎ in cazul cand este descoperitǎ isi retrage capul umflandu-si segmental 1 abdominal, astfel partea anterioarǎ isi- cabreazǎ, in felul acesta face sǎ iasǎ in evidentǎ mari oceli asemǎnǎtori cu niste dandu-I aspectul unei sarpe mic. Pe segmental 8 abdominal are un mic scolus spre deosebire de celelalte omizi de sfingide la care este mare si are un rol defensive.
Rǎspandire: in toatǎ Europa in afarǎ de zonele nordice impǎdurite, prezentǎ si in Romania.
Genul Horisme
Horisme corticata, Treitschke 1835
Perioada de zbor: prezintǎ douǎ generatii:prima generatie tine de la prima jumǎtate a lunii aprilie panǎ in luna iulie; iar a doua generatie incepe la sfarsitul lunii iulie panǎ la sfarsitul lunii septembrie.
Biotop:vǎi si povarnisuri insorite, zǎvoaie, pereti stancosi, podgorii, grǎdini insorite, garduri.
Plante gazda: Clematis vitalba, C.integrifolia, C.alpina.
Ponta si dezvoltarea: femelele in functie de altitudinea in care se aflǎ,depune ouǎle mai ales in luna mai panǎ in luna iulie, larvele se dezvoltǎ repede astfel cǎ la sfarsitul lunii iulie eclozeazǎ fluturii generatiei a doua,femelele depune ponta in luna august panǎ in luna septembrie astfel cǎ la sfarsitul lunii septembrie larvele se impupeazǎ; crisalida petrece iarna sub acest stadium; eclozarea adultilor in primǎvara urmǎtoare.
Rǎspandire: prezentǎ in Europa si in Romǎnia, prezenta legatǎ strans de plantele gazdǎ. Consideratǎ de Koch ca specie alpine.
Genul Operoptera
Operophtera brumata, Linnaeus 1758
Perioada de zbor: din prima jumǎtate a lunii octombrie panǎ la sfarsitul lunii decembrie.
Biotop: pǎduri de foioase, mixte, livezi, tufisuri, grǎdini practice peste tot unde este vegetatie arborescentǎ.
Planta gazda: toti pomii fructiferi ( Mallus sp, Pyrus sp, Prunus sp,) Quercus sp, Betula sp, Prunus spinosa si Crataegus monogyna, practice se hrǎnesc cu frunzele tuturor speciilor de arbori cu frunze cǎzǎtoare. La campie omizile preferǎ mai ales planta Calluma vulgaris.
Ponta si dezvoltarea: femelele depune ouǎle in luna noiembrie panǎ in decembrie in crǎpǎturile scoartei copacilor din care in luna aprilie eclozeazǎ larvele ce-si tes din mǎtase un cuib, prinzand frunzele, de unde ies si se hrǎnesc cu mugurii si frunzele arborilor iar in livezi atacǎ si florile devenind astfel dǎunatoare. In cazul unor dezvoltǎri deosebite se iau mǎsuri radicale, folosinduse diferite insecticide chimice. Isi definesc dezvotarea in luna mai, impupandu-se in sol. De mentionat cǎ femelele au aripile foarte reduse, improprii zborului. Practic adultii apar in iarna.
Rǎspandire: in Europa si in Romania.
Genul: Ligdia
Ligdia adustata, Denis & Schiffermuller 1775
Perioada de zbor: prezintǎ douǎ generatii; prima generatie incepe in prima jumǎtate a lunii aprilie panǎ in luna iulie; iar a doua generatie incepe in prima jumǎtate a lunii iulie panǎ in luna septembrie.
Biotop: pǎduri de amestec, de conifere, parcuri, grǎdini etc.
Plante gazda: Euonymus europea
Ponta si dezvoltarea: femelele depune ouǎle pe planta gazdǎ in perioadele de zbor.
Raspandire: in Europa si in Romania.
Genul Epione
Epione repandaria, Denis & Schiffermuller 1775
Perioada de zbor: prezintǎ douǎ generatii: prima generatie incepe in luna iunie si tine panǎ in prima jumǎtate a lunii august; iar a doua generatie incepe in prima jumǎtate a lunii august si tine panǎ in prima jumǎtate a lunii octombrie.
Biotop: pǎduri si liziere, lunci ,grǎdini.
Ponta si dezvoltarea: femelele in prima generatie depune ponta in luna iunie-iulie, fiind rezultatul larvelor eclozate din ouǎle ce au iernat, apoi femelele in a doua generatie depune ouǎle in zbor la inceputul lui august, astfel cǎ adultii eclozeazǎ in aceeasi lunǎ iar ouǎle depuse ierneazǎ , din ele in urmǎtoarea primavarǎ eclozeazǎ larvele.Impuparea in frunze prinse cu mǎtase.
Rǎspandire: in Europa, in afarǎ de regiunile nordice, de asemenea si in Romania.
Genul: Campea
Campea margaritata, Linnaeus 1761
Perioada de zbor: prezintǎ douǎ generatii; prima generatie incepe in luna mai/iunie si tine panǎ in luna iulie;iar a doua generatie incepe in luna iulie si tine panǎ in luna septembrie/octombrie.
Biotop: pǎduri de foioase, de amestec preferǎ fagetele dumbravi, parcuri.
Plante gazda: Fagus sylvatica, Carpinus betulus, Quercus sp, Betula pendula Salix caprea Prunus spinosa si alte specii de arbori si arbusti de foioase.
Ponta si dezvoltarea: femelele depune ouǎle in luna iunie-iulie in cazul in care prezintǎ douǎ generatii si in luna august-septembrie larva ierneaza continandu-si dezvoltarea in primǎvara urmǎtoare, impupandu-se la suprafata solului.
Rǎspandire: prezentǎ aproape in toatǎ Europa si in Romania.
Fam Noctuidae
Genul Noctua
Noctua pronuba, Linnaeus 1758
Perioada de zbor: incepe in luna mai/iunie si dureazǎ panǎ in luna septembrie/octombrie.
Biotop: prezentǎ peste tot, campuri, grǎdini, parcuri, tufǎrisuri etc.
Plante gazda.:Taraxacumsp, Stellaria media, Rumex sp, Primula sp, Beta vulgaris, Nicotiana tabacum, Vitis vinifera, diferite sp de graminee si alte plante joase cultivate si din flora spontanǎ, puietii din pepiniere pomicole, culturi de flori, pot intra si in sere unde produc daune mari.
Ponta si dezvoltarea: femelele depune ponta ouǎle in luna iulie pe dosul frunzelor. Omizile eclozeazǎ in luna august si intrǎ la iernat spre sfarsitul toamnei, primǎvara urmatoare isi continuǎ dezvoltarea. Se hrǎnesc cu toatǎ frunza, adesea reteazǎ plantele la nivelul solului. Ziua stau ascunse in sol. Se impupeazǎ prin luna mai, fluturii eclozeazǎ, in luna iunie si iulie si zboarǎ panǎ toamna tarziu. Criselida este rosie-brunǎ.
Rǎspandire: sp comunǎ in Europa si in Romania.
Noctua fimbriata, Schreber 1759
Perioda de zbor: incepe in luna iunie si tine panǎ in luna septembrie
Biotop:pante, vǎi, campuri insorite, calde, mixte, grǎdini, parcuri etc
Plante gazda:Primula vulgaris, Rubus ideaus,R. fructicosus, Viola odorata, Lamium sp., Crataegus monogyna, Prunus spinosa, Betula pendula, Salix sp., Vitis vinifera si alte plante cu frunze joase,diverse spp. de legume
Ponta si dezvoltarea: femelele depune ponta la sfarsitul verii, omizile inainte de-a intra la iernat cautǎ plante joase,in primǎvara se hrǎnesc cu frunze de arbori si arbusti de foioase.Alti autori considerǎ cǎ femelele depune ouǎle intre frunzele arborilor iar omizile pentru a ierna coboarǎ pe sol ( C a r t e r si colab., 1988). In luna mai se impupeazǎ la suprafata solului.Fluturii eclozeazǎ in luna iunie panǎ in luna iulie
Rǎspandirea: Sp. Comunǎ in Europa si in Romania
Subgenul Melanchra Hubner, 1820
Malanchra persicariae persicariae Linnaeus, 1761
Perioda de zbor: incepe in luna mai si se terminǎ in luna august
Biotop: Peste tot, margini de pǎduri, poieni, lunci, campuri cu vegetaǎie spontana si cultivate, grǎdini, parloage s.a.
Plante gazda: Urtica sp., Anemone sp., Rubus idaeus, Sanpucus sp., S. nigra, Plantago sp ., Nicotina tabacum, diverse legume si alte sp. de plante joase, arbusti si arbori de foioase.
Ponta si dezvoltarea: Ouǎle depuse izolat sau in grupe pe planta gazdǎ in luna iunie, omizile eclozeazǎ dupǎ 7-8 zile si se hrǎnesc noaptea , ziua stau nemiscate sub frunze pe planta gazdǎ.Impuparea in sol intr-un cocon in care se aflǎ crisalida ce va ierna. Din ea eclozeazǎ fluturi in luna mai. Crisalida rosie brunǎ ierneazǎ. Larvele frecvent sunt associate cu cele de M. brasicae, M.olaracea, M.pisi, Discestra trifolii.
Rǎspandirea: comunǎ in Europa si in Romania .
Relatia om-fluture: Participǎ adesea la defolierea culturilor de sfecla de zahar si chiar vǎrzoase, in compania altor speccii de omizi. Mǎsurile de reducerea populatiilor sub PED sunt aceleasi ca la M.brasicae. Larvele creazǎ probleme deosebite numai in companie cu alte sp.
Diversitatea entomofaunei nocturne
In perioada iulie - septembrie la cantonul silvic Cetatuia a functionat o capcana luminoasa la care s-a colectat entomofauna nocturna. Zona este caracterizata printr-o padure de amestec de carpen, tei, stejar, iar situarea in apropiere a unui iaz permite pastrarea unui climat mai umed. De asemenea, langa canton se afla o plantatie de lucerna pentru uz silvic (ca furaj), ceea ce atrage o serie intreaga de insecte, mai ales in perioada de inflorire. Capcana a fost folosita de asemenea si in luna noiembrie, insa din cauza temperaturilor scazute nu s-au colectat decat cateva exemplare de coleoptere, motiv pentru care probele au fost considerate nesemnificative din punct de vedere cantitativ.
Entomofauna nocturna a fost dominata de grupele de insecte holometabole din ordinele Lepidoptera, Diptera, Coleoptera, care ating efectivele cele mai mari - peste 2400 exemplare de lepidoptere in iunle, peste 600 in august, peste 1000 exemplare de nematocere in iulie si peste 3000 in septembrie. Toate aceste date vin sa sugereze ca zona analizata se caracterizeaza printr-o biodiversitate taxonomica extrem de bogata.
Efectivele grupelor taxonomice din entomofauna nocturna la Cetatuia, in intervalul iulie - septembrie 2006
|
Grup taxonomic |
Numar exemplare colectate |
||
|
iulie |
august |
septembrie |
|
|
Homoptera (Ciccadina, Aphidina, Psyllina) | |||
|
Heteroptera | |||
|
Coleoptera | |||
|
Neuroptera | |||
|
Lepidoptera | |||
|
Diptera - Nematocera | |||
|
Diptera - Brachicera | |||
|
Hymenoptera | |||
|
Total | |||
In luna iulie, probele au fost luate aproape in fiecare noapte, in intervalul 1 - 7.07. 2006. Analizand procentual ponderea diferitelor grupe taxonomice, se constata ca dominante in aceasta perioada au fost lepidopterele, cu 41%, urmate de nematocere si coleoptere cu cate 15% fiecare. Restul grupelor taxonomice inregistreaza valori sub 10%.
Analizand evolutia efectivelor diferitelor grupe taxonomice, se constata ca cele mai bogate capturi au fost la inceputul intervalului, cand zona a fost afectata de un val puternic de caldura iar noptile au fost neobisnuit de calduroase. Ulterior, toate efectivele inregistreaza descresteri numerice, cu exceptia dipterelor brahicere, care manifesta o tendinta de crestere exact in perioada in care celelalte grupe taxonomice au efectivele minime. Acest aspect poate fi pus pe seama preferintelor ecologice ale brahicerelor, iubitoare de umiditate.
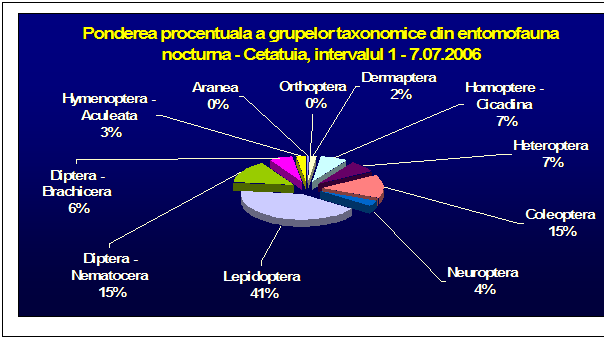
In luna august, intervalul analizat a fost mai extins - 14 - 24 august. Din punct de vedere cantitativ, grupa dominanta si in acest caz a fost cea a lepidopterelor (43%), urmata tot de nematocere si coleoptere, dar procentele acestor doua grupe sunt sensibil mai mari comparativ cu perioada precedenta - 21% respectiv 14%. O pondere semnificativa, de 10%, au in aceasta perioada si heteropterele. Celelalte grupe au ponderi situate sub aceasta din urma valoare.
Din punct de vedere al evolutiei
efectivelor, se constata un fapt interesant: efectivele de lepidopterele
si dipterele nematocere variaza in acelasi ritm, practic avand
aceleasi intervale de maxim (14 - 17.08, 18 - 20.08, 21 - 22.08) si
de minim (17 - 18.08, 20 - 21.08). O evolutie interesanta au si efectivele de coleoptere,
insa in acest caz, maximele efectivelor de coleoptere sunt concomitente cu
perioade in care efectivele de nematocere au valorile minime. In ceea ce
priveste comparatia cu efectivele de lepidoptere, in acest caz se
constata o situatie similara cu exceptia ultimei perioade -
21 - 22 august - cand ambele grupe taxonomice inregistreaza maxime.
Heteropterele au si ele o evolutie asemanatoare cu cea a
efectivelor de 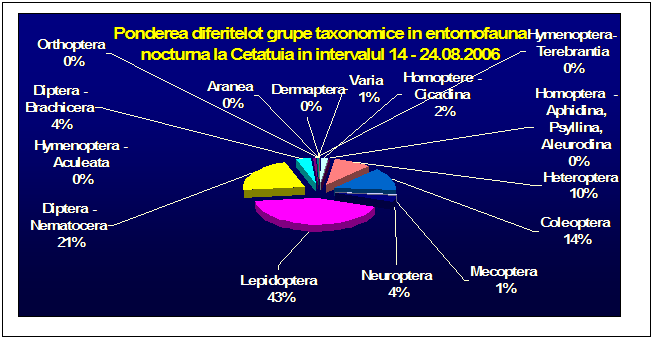 coleoptere, curbele fiind insa putin
defazate in acest caz.
coleoptere, curbele fiind insa putin
defazate in acest caz.
In luna septembrie, perioada de colectare s-a situat in intervalul 26 - 30.09. Comparativ cu lunile de vara, in aceasta perioada se constata o schimbare radicala a structurii entomofaunei nocturne. Dominante devin in aceasta perioada dipterele nematocere, kar efectivele lor depasesc cu mult efectivele tutoror celorlalte grupe taxonomice - 90%. Dintre celelalte grupe taxonomice, dipterele brahicere, lepidopterele si homopterele inregistreaza fiecare cate 2%.
Analiza variatiei efectivelor diferitelor grupe indica un maxim pentru brahicere, coleoptere, lepidoptere, himenoptere, homoptere in prima parte a intervalului (27.09.2006), in vreme ce maximul pentru nematocere se inregistreaza spre sfarsitul intervalului. In aceeasi perioada mai inregistreaza maxime si lepidopterele, homopterele si heteropterele.
In acest caz, schimbarea structurii entomofaunei este datorata evolutiei anotimpurilor si schimbarii tipului de entomofauna. De asemenea, marea abundenta de nematocere poate fi pusa direct in lefatura cu prezenta unei zone umede in imediata vecinatate, cu vegetatia caracteristica.
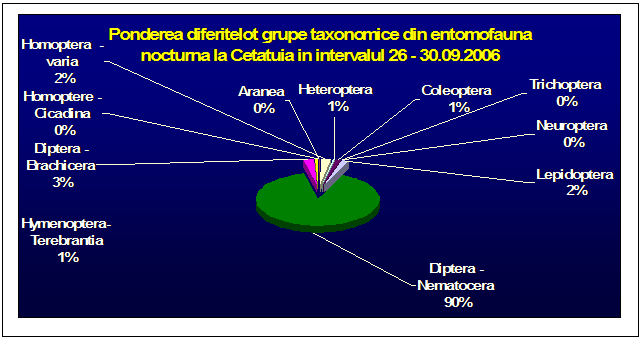
Analizand tendinta evolutiei efectivelor de insecte din entomofauna nocturna pentru intregul interval de studiu se constata relativ usor ca patru grupe taxonomice caracterizeaza ceea ce putem denumi entomofauna tipic de vara: lepidopterele, coleopterele, heteropterele si neuropterele. Cu toate ca inregistreaza efective diferite, toate aceste grupe au o tendinta clara de scadere a efectivelor, evidentiata extrem de bine pe un grafic semilogaritmic. Pe de alta parte, dipterele, atat nematocere cat si brahicere, inregistreaza o perioada de diminuare a efectivelor in luna august, pentru ca sa devina grupe dominante pentru entomofauna de toamna. Tendinta este extrem de evidenta la nematocere, dar poate fi evidentiata de asemenea la brahicere, chiar daca in acest caz efectivele sunt mult mai reduse.
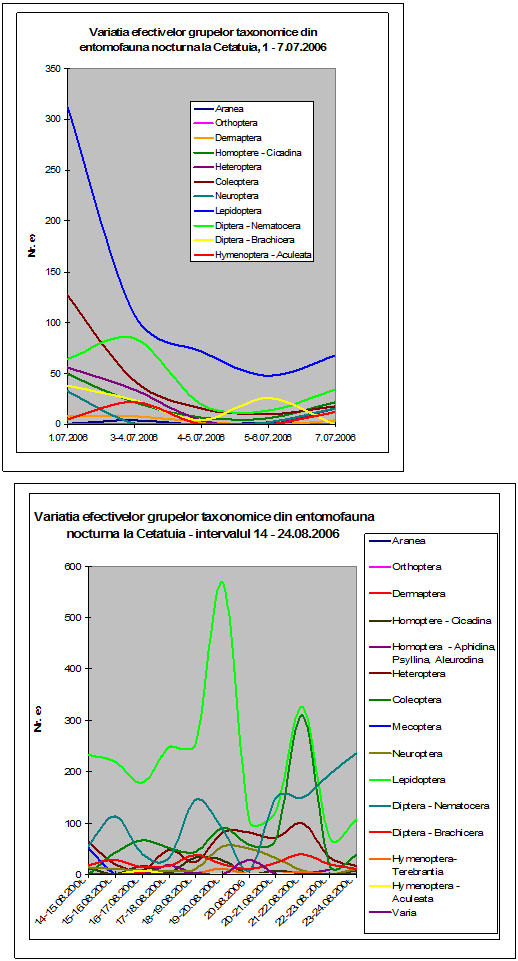
Structura faunei de lepidoptere nocturne
Cu toate ca studiul entomofaunei nocturne s-a limitat la colectari la lumina efectuate doar in zona Cetatuia si nu au fost facute colectari in cele mai interesante habitate ale PNMM decat ocazional pentru acest tip de entomofauna, totusi au fost identificate 130 de specii de lepidoptere nocturne. Trebuie mentionat ca determinarea nu a fost incheiata si mai ales pentru microlepidoptere datele obtinute sunt doar partiale.
Cu toate acestea, spectrul taxonomic al lepidopterelor nocturne arata unele date interesante. Au fost identificate unele specii deosebite din punct de vedere zoogeografic - mentionate de altfel din alte zone de studiile anterioare.
Fauna de lepidoptere nocturne este dominata astfel de speciile familiei Noctuidae, care ocupa 39% din totalul speciilor, fiind urmate de Geometridae, cu 37%. Celelalte familii sunt reprezentate prin specii putine - Sphingidae 5%, Notodontidae 5%, Arctiide 4% etc, structura care corespunde in mare structurii taxonomice a macrolepidopterelor nocturne de pe teritoriul Dobrogei.
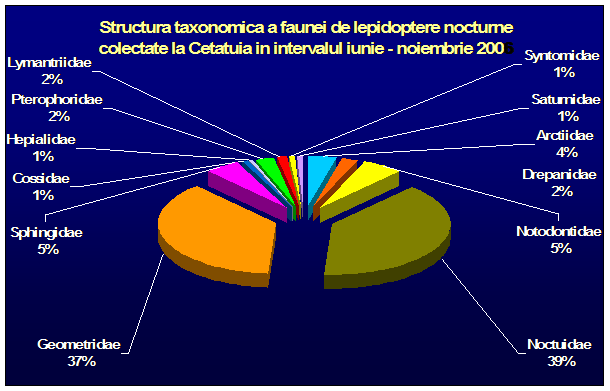
Analizand variatia numarului de specii din cele doua familii identificate in special in zona Cetatuia, se constata ca pentru ambele familii luna august marcheaza un maxim urmat de scaderea numarului de taxoni in perioada lunilor de toamna. Cel mai favorabil interval pentru analiza in profunzime a structurii taxonomice a faunei de lepidoptere nocturne apare astfel luna august. Cu toate acestea, o serie de specii au perioade de zbor cuprinse in intervalul mai - iulie si se impune cu necesitate extinderea studiului si in lunile respective pentru obtinerea unei priviri de ansamblu asupra entomofaunei de lepidoptere nocturne.
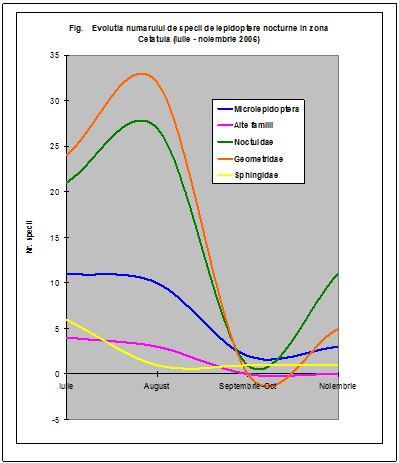
|
iulie |
august |
Septembrie-oct |
noiembrie |
|
|
Cossidae | ||||
|
Phragmatecia castanea Hb. | ||||
|
Hepialidae | ||||
|
Hepialus sylvinus L. | ||||
|
Pterophoridae | ||||
|
Pterophorus pentadactyla L. | ||||
|
Capperia britanniodactyla | ||||
|
Alucita hexadactyla | ||||
|
Gracillariidae | ||||
|
Cameraria ohridella D. et D.(Cetatuia, pe castan ) | ||||
|
Phylonorycter robiniella Clemens | ||||
|
Pyralidae | ||||
|
Hypsopygia costalis Fabr. | ||||
|
Pyrausta aurata Scop. | ||||
|
Pyrausta purpuralis L. | ||||
|
Pleurota ruralis Scop. | ||||
|
Paraponyx stagnata D. | ||||
|
Onocera semirubella Scop. | ||||
|
Eurrhypara hortulata L. | ||||
|
Crambus pratella L. (f. com) | ||||
|
Pyrausta aurata Scop.(f. com) | ||||
|
Ostrinia nubilalis Hb. | ||||
|
Lymantriidae | ||||
|
Lymantria disparL. (f. melanica) | ||||
|
Arctornis l-nigrum Mull. | ||||
|
Arctiidae | ||||
|
Miltochrista miniata Forst. | ||||
|
Oenistis quadra L. | ||||
|
Phragmatobia fuliginosa L. | ||||
|
Lithosia lutarella L. | ||||
|
Panaxia quadripunctaria Poda | ||||
|
Drepanidae | ||||
|
Cilix glaucata scop. | ||||
|
Drepana harpagula Esp. | ||||
|
Drepana binaria Hufn. | ||||
|
Axiidae | ||||
|
Habrosyne pyritoides Hfn. | ||||
|
Notodontidae | ||||
|
Pterostoma palpina Cl. | ||||
|
Habrosyne derasa L. | ||||
|
Clostera anastomosis L. | ||||
|
Odonestis pruni L. | ||||
|
Pygaera anastomosis L. | ||||
|
Spatalia argentina D. et S. | ||||
|
Syntomidae | ||||
|
Dysauxes ancilla l. | ||||
|
Noctuidae | ||||
|
Abrostola triplasia L. | ||||
|
Aedia leucomelas L. | ||||
|
Amphipyra livida D. et S. | ||||
|
Amphypyra pyramidea L. | ||||
|
Apamea monoglypha Hfn. | ||||
|
Asteroscopus sphinx Hfn. |
+ necitat de rakosy |
|||
|
Agrochola circellaris Hfn. | ||||
|
Conistra veronicae Hb. | ||||
|
Autographa confusaHb. | ||||
|
Autographa gamma L. | ||||
|
Calophaena opalina | ||||
|
Catocala elocata Esp. | ||||
|
Catocala nupta L. | ||||
|
Colobochyla salicalis D. et S. (?) | ||||
|
Cosmia pyralina D. et S. | ||||
|
Cucullia fraudatrix Ev. | ||||
|
Deltote bankiana Fabr. | ||||
|
Diachrysia chrysitis L. | ||||
|
Diachrysia chryson Esp. | ||||
|
Elaphria venustula Hb. | ||||
|
Erastria trabealis Scop. | ||||
|
Eucarta virgo Treit. | ||||
|
Gonospileia glyphica L. | ||||
|
Helicoverpa armigera Hb. | ||||
|
Helicoverpa peltigera D. et S. | ||||
|
Heliothis cardui Hb. (V. Ghiunaltu) |
necitat de rakosy | |||
|
Heliothis maritima bulgarica Draut. | ||||
|
Hylophila prasinana | ||||
|
Hypena proboscidalis l. | ||||
|
Lygephila craccae D. et. S. | ||||
|
Melanchra persicariae L. | ||||
|
Mycteroplus puniceago Bsdv. | ||||
|
Mythimna congrua Hb. | ||||
|
Naena typica | ||||
|
Noctua fimbriata Schreber | ||||
|
Noctua pronuba L. |
pasul Teilor |
Pietrosul II |
|
|
|
Prodotis stolida Fabr. | ||||
|
Protoschinia scutosa D. et S. | ||||
|
Pseudeustrotia candidula D. et S. | ||||
|
Pseudoips fagana fabr. | ||||
|
Pyrrhia umbra Hfn. | ||||
|
Rusina ferruginea Esp. | ||||
|
Scoliopterix libatrix L. | ||||
|
Sideritis conigera D. et S. (f. com) | ||||
|
Tyta luctosa D. et S. | ||||
|
Toxocampa craccae F. (?) | ||||
|
Trachea atriplicis L. | ||||
|
Ulochlaena hirta Hb. |
necitat de rakosy |
|||
|
Viminia rumicis L. | ||||
|
Xestia c-nigrum L. | ||||
|
Xylina vetsusta L. (Nifon) | ||||
|
Geometridae | ||||
|
Operoptera brumata L. |
+ necitat de rakosy |
|||
|
Anaitis plagiata L. | ||||
|
Anticlea derivata D. et S. | ||||
|
Aspilates gilvaria D. et S. | ||||
|
Boarmia rhomboidaria D. et S. | ||||
|
Boarmia selenaria Schiff. | ||||
|
Calothysanis amata L. | ||||
|
Campaea margaritata L. | ||||
|
Camptograma bilineata | ||||
|
Chlorissa viridata L. | ||||
|
Cidaria ocellata L. | ||||
|
Cosmorhoe ocellata L. | ||||
|
Cosymbia orbicularis | ||||
|
Cosymbia punctaria L. | ||||
|
Cosymbia pupillaria Hb. | ||||
|
Cyclophora albipunctata Hb. | ||||
|
Cyclophora annulata Sch. | ||||
|
Ematurga atomaria L. | ||||
|
Endrosis sarcetella (de verificat) - 27.09. | ||||
|
Ennomos erosaria D. et S. | ||||
|
Ennomos quercinaria Hfn. | ||||
|
Epione repandaria Hfn. | ||||
|
Euchloris smaragdaria fabr. | ||||
|
Euphitecia centaureata D. et S. | ||||
|
Euphitecia extraversaria H.S.? | ||||
|
Euphitecia linariata F. | ||||
|
Euphitecia pyreneata Mab. | ||||
|
Hemistola chrysoprasaria Esp. | ||||
|
Hipparchus papilionaria L. | ||||
|
Horisme corticata Treit. | ||||
|
Horisme terasata | ||||
|
Idaea muricata Hfn. | ||||
|
Ligidia adustata D. et S. | ||||
|
Lomaspilis marginata L. | ||||
|
Lythria purpuraria L. | ||||
|
Mesoleuca albicillata L. | ||||
|
Perizoma lugdunaria H.S. | ||||
|
Plagodis dolabraria L. | ||||
|
Rhodostrophia calabra Petagna | ||||
|
Scopula immorata L. | ||||
|
Scopula rubiginata Hfn. (?) | ||||
|
Selenia tetralunaria Hfn. | ||||
|
Semiothisa clathrata L. | ||||
|
Semiothisa notata L. | ||||
|
Sterrha aversata L. | ||||
|
Sterrha ochrata Scop. | ||||
|
Tephrina arenacearia Hb. | ||||
|
Thalera fimbrialis Scop. | ||||
|
Sphingidae | ||||
|
Macroglossum stellatarum L. | ||||
|
Laothoe populi L. | ||||
|
Marumba quercus L. | ||||
|
Deilephila elpenor L. | ||||
|
Celerio galii Rott. (larva) | ||||
|
Celerio euphorbiae L.(larva) | ||||
|
Fam. Saturnidae | ||||
|
Saturnia pyri L. (larva) |
Botnariuc N., Vadineanu A., 1982 - Ecologie. Ed.Did.si Ped. Bucuresti.
Ciochia V., Stanca Cristina, 2000 - catalogul colectiei didactice de lepidoptere cu elemente de biologie a Universitatii "Lucian Blaga" din Sibiu, Ed Pentru Viata, Brasov.
Cotet P., 1968 - Dobrogea de Sud - geneza si evolutie, in Studii geografice asupra Dobrogii, Bucuresti: 51-58.
Donita N. et colab., 1882 - Vegetatia Romaniei, Ed. Tehnica Agricola, Bucuresti.
Donita N., Ivan Doina, Chifu Th., Mititelu D., Coldea Ghe., 1992 - Vegetatia Romaniei, Editura Tehnica agricola, Bucuresti.
Dragomirescu L., - Biostatistica pentru incepatori, Editura Constelatii, Bucuresti, 1998.
Fira Valeria, Nastasescu Maria, 1977 - Zoologia nevertebratelor, Ed.Did.si Ped. Bucuresti
Gomoiu M.-T., Skolka M., 2000 - Ecologie. Metodologii pentru studii ecologice, Ovidius University Press, Constanta
Grasse P. (red), 1951 - Ordre des Lepidopteres in Traité de zoologie, Masson et Co. Ed., Paris. p 174-448.
Higgins L., Riley N., 1975 - Guide des Papillons d"Europe, Ed. Delachaux et Niestle, Neuchatel - Paris.
Ionescu A., Lacatusu Matilda, 1971 - Entomologie, ed.Did. si Ped. Bucuresti
Ivan Doina, 1979 - Fitocenologie si vegetatia Romaniei, Editura didactica si pedagogica Bucuresti.
Jaba Elisabeta, 1996 - Statistica, Ed. Sedcom Libris, Iasi.
Koch M., 1991 - Schmetterlinge, Newmann Verlag
Krebs
CJ., 1989, - Ecological methodology, Harper Collins Publishers,
Leraut P., 1980 - Liste systematique et synonymique des lepidopteres de France, Belgique et Corse - Abwanar.Bul.Soc.Ent.Fr. Suppl, Paris.
Magurran
A.E.: 1987 - Ecological diversity andits measurements, Princeton University
Press.
Niculescu E.V., Konig Fr., 1970 - Fauna Rom Vol 11 fasc 10 Lepidoptera - Parte Generala. Ed.Acad. Bucuresti.
Popescu-Gorj A., 1964 - Catalogue de la collection de Lepidopteres "Prof.A.Ostrogovich" du Museum d`Histoire Naturelle "Gr.Antipa" Bucharest.
Popovici I., 1984 - Podisul Dobrogei si Delta Dunarii, Ed. Stiintifica si Enciclopedica, Bucuresti.
Radu Gh. Vasile, 1967 - Zoologia nevertebratelor, Editura didactica si pedagogica Bucuresti.
Radu V.Gh., Radu Varvara, 1967 - Zologia nevertebratelor Vol.II, Ed.Did.si Ped., Bucuresti
Sokal R. R., Sneath P.M., 1963 - Principles of numerical Taxonomy, W.F. Fresman and Co., San Francisco and London
Stanoiu M., Bobarnac B., Copacescu S, 1977 - Fluturii din Romania, Ed.Scrisul Romanesc, Craiova.
Stugren
B. 1994 - Ecologie generala, Editura Sarmis,
Stugren B., 1975 - Ecologie Generala. Ed. Did.si Ped. Bucuresti.
Stugren B., 1982 - Bazele ecologiei generale. Ed. St. Enc. Bucuresti.
Stugren B., Killyen H.G., 1975 - Ecologie. Ed. Did.si Ped. Bucuresti.
X X X - Parcul National Muntii Macin, https://www.parcmacin.ro/
ANEXA
|
Noctuidae |
Orig. Zoog. |
Nr.gen. |
Cum ierneaza? |
Ecologie |
||||
|
Abrostola triplasia |
euroasiatica |
nimfa |
mezofila |
|||||
|
Aedia leucomelas |
Euroasiatica subtropicala |
nimfa |
mezotermofila |
|||||
|
Amphypira livida |
euroasiatica |
ou |
mezotermofila |
|||||
|
Amphypira pyramidea |
euroasiatica |
ou |
Mezofila |
|||||
|
Apamea monoglypha |
euroasiatica |
larva |
Eurioica |
|||||
|
Asteroscopus sphinx |
euroasiatica |
ou |
Mezofila |
|||||
|
Agrochola circellaris |
Euroasiatica |
ou |
Mezofila |
|||||
|
Conistra veronicae |
mediteraneeana |
Adultul(hiberneaza) |
Mezotermofila |
|||||
|
Autographa confusa |
euroasiatica |
larva |
Eurioica(sp ruderala) |
|||||
|
Autographa gamma |
Euroasiatica |
Larva-pupa-adult |
eurioica |
|||||
|
Calophaena opalina |
mediteraneena |
larva |
Xerotermofila |
|||||
|
Catocala elocata |
IRPM |
ou |
Termofila |
|||||
|
Catocala nupta |
euroasiatica |
ou |
Mezofila |
|||||
|
Colobochyla salicalis |
euroasiatica |
pupa |
mezohigrofila |
|||||
|
Cosmya pyralina |
Euroasiatica |
ou |
Mezohigrofila |
|||||
|
Cucullia fraudatrix |
euroasiatica |
pupa |
Mezoxerotermofila |
|||||
|
Deltote bankiana |
euroasiatica |
pupa |
Mezohigrofila |
|||||
|
Diachrysia chrysitis |
Euroasiatica |
larva |
mezofila |
|||||
|
Diachrysia chryson |
Euroasiatica |
larva |
mezohigrofila |
|||||
|
Elaphria venustula |
euroasiatica |
pupa |
Mezotermofila |
|||||
|
Erastria trabealis |
euroasiatica |
pupa |
mezotermofila |
|||||
|
Eucarta virgo |
euroasiatica |
pupa |
Mezotermohigrofila |
|||||
|
Gonospileia glyphica |
euroasiatica |
pupa |
Mezotermofila |
|||||
|
Helicoverpa armigera |
Tropicala-cosmopolita |
pupa |
Termofila(ruderala) |
|||||
|
Helicoverpa peltigera |
subtropicala |
nimfa |
Termofila(ruderala) |
|||||
|
Schinia cardui |
IRPM |
pupa |
Xerotermofila |
|||||
|
Heliothis maritima bulgarica |
euroasiatica |
pupa |
Xerotermofila |
|||||
|
Hylophila prasinana |
euroasiatica |
pupa |
Mezofila |
|||||
|
Hypena proboscidalis |
euroasiatica |
omida |
Mezohigrofila |
|||||
|
Lygephila craccae |
euroasiatica |
pupa |
Mezoxerotermofila |
|||||
|
Melanchra persicariae |
euroasiatica |
pupa |
Mezohigrofila |
|||||
|
Mycteroplus puniceago |
IRPM |
ou |
Xerotermofila |
|||||
|
Mythimma congrua |
IRPM |
larva |
Termohigrofila |
|||||
|
Naena typica |
euroasiatica |
omizile |
Mezofila |
|||||
|
Noctua fimbriata |
mediteraneeana |
omida |
Mezofila |
|||||
|
Noctua pronuba |
euroasiatica |
omida |
Mezofila |
|||||
|
Prodotis stolida |
IRPM |
pupa |
Xerotermofila |
|||||
|
Protoschinia scutosa |
holartica |
pupa |
Xerotermofila |
|||||
|
Pseudeustrotia candidula |
euroasiatica |
pupa |
Mezohigrofila |
|||||
|
Pseudoips fagana | ||||||||
|
Pyrrhia umbra |
holartica |
pupa |
Mezofila |
|||||
|
Russina ferruginea |
euroasiatica |
larva |
Mezofila |
|||||
|
Scoliopterix libatrix |
holartica |
adultul |
mezofila |
|||||
|
Sideritis conigera |
euroasiatica |
omida |
Mezofila | |||||
|
Tyta luctosa |
euroasiatica |
Pupa |
Xerotermofila | |||||
|
Toxocampa craccae | ||||||||
|
Trachea atriplicis |
euroasiatica |
pupa |
Mezofila | |||||
|
Ulochlaena hirta |
mediteraneeana |
ou |
Xerotermofila | |||||
|
Vimina rumicis |
euroasiatica |
pupa |
Mezofila | |||||
|
Xestia c-nigrum |
cosmopolita |
larva |
eurioic | |||||
|
Xylina vetsusta | ||||||||
|
Geometridae | ||||||||
|
Operoptera brumata |
Eurosiberiana |
ou |
Criofil | |||||
|
Anaitis plagiata |
larva |
Termofila | ||||||
|
Anticlea derivata |
pupa |
Termohigrofil | ||||||
|
Aspilates gilvaria |
eurosiberiana |
larva |
Termofila | |||||
|
Boarmia rhomboidaria |
larva |
Higrofil | ||||||
|
Boarmia selenaria |
pupa |
Mezohigrofil | ||||||
|
Calothysanis amata |
larva |
Mezofila | ||||||
|
Campaea margaritata |
euroasiatica |
Larva |
Higrofila | |||||
|
Camptograma bilineata | ||||||||
|
Chlorissa viridata |
euroasiatica |
pupa |
mezohigrofil | |||||
|
Cidaria ocellata |
larva |
Mezohigrofil | ||||||
|
Cosmorhoe ocellata | ||||||||
|
Cosymbia orbicularis |
pupa |
Higrofila | ||||||
|
Cosymbia punctaria |
pupa |
Higrofila | ||||||
|
Cosymbia pupillaria |
pupa |
Mezotermofil | ||||||
|
Cyclophora albipuncta |
euroasiatica |
pupa |
Mezohigrofil | |||||
|
Cyclophora annulata |
euroasiatica |
pupa |
Mezohigrofil | |||||
|
Ematurga atomaria |
pupa |
Mezohigrofil | ||||||
|
Endrossis sarcetella | ||||||||
|
Ennomos erosaria |
eurosiberiana |
ou |
Mezofil | |||||
|
Ennomos quercinaria |
ou |
Mezohigrofil | ||||||
|
Epione repandaria |
euroasiatica |
Ou |
Higrofil | |||||
|
Euchloris smaragdaria |
larva |
mezohigrofil | ||||||
|
Euphitecia centaureata |
euroasiatica |
pupa |
Mezofila | |||||
|
Euphitecia extraversaria |
eurosiberiana |
pupa |
Termifila | |||||
|
Euphitecia linariata |
eusiberiana |
pupa |
Mezofil | |||||
|
Euphitecia pyreneata |
pupa |
Mezofil | ||||||
|
Hemistola chrysoprasaria |
larva |
Mezofil | ||||||
|
Hipparchus papilionaria |
larva |
Mezofil | ||||||
|
Horisme corticata |
euroasiatica |
pupa |
Termofila | |||||
|
Horisme terasata |
euroasiatica |
pupa |
Higrofil | |||||
|
Idaea muricata |
euroasiatica |
larva |
Mezohigrofil | |||||
|
Ligidia adustata |
eurosiberiana |
pupa |
Mezohigrofil | |||||
|
Lomaspilis marginata |
pupa |
Higrofil | ||||||
|
Lythria purpuraria |
pupa |
Higrofil | ||||||
|
Mesoleuca albicillata |
pupa |
Mezohigrofil | ||||||
|
Perizoma lugdunaria |
eurosiberiana |
pupa |
termofila | |||||
|
Plagodis dolabraria |
eurosiberiana |
pupa |
Mezotermofil | |||||
|
Rhodostrophia calabra |
larva |
mezotermofil | ||||||
|
Scopula immorata |
euroasiatica |
larva |
Mezotermofil | |||||
|
Scopula rubiginata |
euroasiatica |
larva |
Mezotermofil | |||||
|
Selenia tetralunaria |
eurosiberiana |
pupa |
Mezohigrofil | |||||
|
Semiothisa clathrata |
euroasiatica |
pupa |
Higrofil | |||||
|
Semiothisa notata |
eurosiberiana |
Pupa |
Higrofil | |||||
|
Sterrha aversata |
larva |
mezohigrofil | ||||||
|
Sterha ochrata |
larva |
Termofila | ||||||
|
Tephrina arenacearia |
eurosiberiana | |||||||
|
Thalera fimbrialis |
larva |
Mezohigrofil | ||||||
|
Sphingidae | ||||||||
|
Macroglosum stellatarum |
Euroasiatica |
nimfa |
Mezohigrofila | |||||
|
Laothoe populi |
eurosiberiana |
pupa |
Mezohigrofil | |||||
|
Marumba quercus |
IRPM |
nimfa |
Mezotermofil | |||||
|
Deilephila elepenor |
euroasiatica |
nimfa |
Mezohigrofila | |||||
|
Celerio galii |
holartic |
nimfa |
Mezohigrofila | |||||
|
Celerio euphorbiae |
euroasiatica |
larva |
mezohigrofila | |||||
|
Lymantriidae | ||||||||
|
Lymantria dispar |
holartica |
nimfa |
Mezofila | |||||
|
Arctornis nigrum |
euroasiatica |
larva |
mezohigrofil | |||||
|
Arctiidae | ||||||||
|
Miltochrista miniata |
euroasiatica |
pupa |
Mezohigrofila | |||||
|
Oenistis quadra | ||||||||
|
Phragmatobia fuliginosa |
euroasiatica |
larva |
Mezohigrofila | |||||
|
Lithosia lutarella |
euroasiatica |
larva |
termofila | |||||
|
Panaxia quadripunctaria |
eurosiberiana |
larva |
Hidrofil | |||||
|
Drepanidae | ||||||||
|
Cilix glaucata | ||||||||
|
Drepana harpagula |
pupa |
Mezohigrofil | ||||||
|
Drepana binaria |
eurosiberiana |
pupa |
Mezotermofil | |||||
|
Axiidae | ||||||||
|
Habrosyne pyritoides |
eurosiberiana | |||||||
|
Notodontidae | ||||||||
|
Pterostoma palpina |
eurosiberiana |
larva |
Mezohigrofila | |||||
|
Habrosyne derasa | ||||||||
|
Clostera anastomosia |
euroasiatica |
Mezohigrofila | ||||||
|
Odenestis pruni |
larva |
mezohigrofil | ||||||
|
Politica de confidentialitate |
| Copyright ©
2025 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |