MORFOLOGIE BACTERIANA
Bacteriile sunt vietuitoare unicelulare
Celula este protejata de un perete (membrana) cu o structura rigida
care confera fiecarei specii o forma caracteristica.
Se disting patru tipuri morfologice principale:
Multiplicarea bacteriilor - prin sciziparitate, aparatul nuclear se divide pe cale amitotica (prin replicarea filamentului de ADN)
Exceptional, anumite grupe de bacterii se pot multiplica si pe alte cai
(inmugurire,fragmentare etc.).
Bacteriile sunt lipsite de pigmenti fotosintetizanti
* cu exceptia catorva grupe de bacterii
Energia necesara activitatii vitale si-o procura prin reactii chimice exergonice.
vietuitoare strict chemotrofe (chemosintetizante).
poseda un bogat echipament enzimatic, care le permite sa utilizeze, in metabolismul plastic si energetic, componente organice si anorganice foarte variate.
Lumea bacteriana poate fi impartita in doua categorii:
Caracterele morfologice proprii unor specii sau grupe de bacterii au o
Servesc ca importante criterii taxonomice
Utilitate practica esentiala in lucrarile curente de diagnostic bacteriologic.
FORMA
Patru tipuri morfologice:
Cocii
au o forma sferoidala, aparent rotunda, uneori usor ovalara sau chiar neregulata.
Unele specii de coci au o forma particulara caracteristica;
Streptococcus pneumoniae
(pneumococ) - celulele bacteriene
au forma ovalara lanceolata
Neisseria gonorrhoeae
(gonococ) o forma ovalara reniforma.
Bacilii
au forma cilindrica, mai mult sau mai putin alungita, asemanatoare unui bastonas.
La unele specii forma bacilara este usor aplatizata (Erysipelothrix insidiosa)
Numeroase specii de bacili prezinta obisnuit aspecte de polimorfism:
Intre bacili exista deosebiri in privinta grosimii si lungimii sau a formei pe care o au capetele acestora.
La unele specii formele bacilare sunt subtiri, fine, de regula rectilinii (E. insidiosa),
la altele pot fi usor incurbate (Mycobacterium tuberculosis);
La majoritatea speciilor capetele bacililor sunt usor rotunjite
B. anthracis capetele sunt insa drepte.
La speciile din genul Fusobacterium celula se subtiaza spre capete dand bacteriilor un aspect fusiform.
Vibrionii
au forma alungita si incurbata, uneori cu ondulatii neregulate.
Speciile din genul Vibrio au obisnuit o forma incurbata, asemanatoare cu o virgula sau cu un corn.
Spirochetele
au aspect filiform, filamentele fiind foarte subtiri si lungi, cu numeroase ondulatii sub forma de spire regulate (asemanatoare unui tirbuson).
Aspectul spirelor poate fi diferit:
spire numeroase (cateva zeci) foarte stranse si regulate;
spire mai largi si mai putin regulate.
Aspectul spirelor poate fi diferit:
speciile din genul Leptospira au spire numeroase (cateva zeci) foarte stranse si regulate;
speciile din genul Borrelia au spire mai largi si mai putin regulate.
MOD DE GRUPARE
celulele rezultate din diviziune se separa, raspandindu-se neuniform in mediu.
La unele bacterii celulele raman insa atasate una de alta, de o maniera care duce la formarea de grupari caracteristice
modul de grupare constituie la unele specii un important criteriu taxonomic
Cauzele care determina constituirea de grupari caracteristice sunt multiple
Felul in care se realizeaza diviziunea celulelor bacteriene - la unele specii totdeauna pe anumite planuri si directii determinate genetic
Mentinerea lor atasata se realizeaza in majoritatea cazurilor printr-o substanta vascoasa, secretata numai pe anumite zone de la suprafata bacteriei, determinate genetic
Cele mai caracteristice modalitati de grupare sunt intalnite la coci
Diplococii (diplos = dublu). Sunt coci grupati cate doi.
fiecare celula bacteriana se divide totdeauna pe un singur plan, iar celulele fiice raman in continuare atasate, realizandu-se astfel grupari de cate
doua celule sau
inlantuiri scurte de asemenea diplococi.
La unele specii, aranjamentul fata in fata a celor doi coci se face de o maniera foarte caracteristica.
Str. pneumoniae (pneumococul) are o forma lanceolata si se dispune totdeauna cu partea ascutita fata in fata,
Neisseria gonorrhoeae (gonococul) are forma reniforma si se dispune totdeauna cu partea reniforma fata in fata.
Streptococii (streptos = lant) - coci dispusi intr-o inlantuire liniara, asemanatoare unui sirag de margele
Gruparea in lant este determinata de faptul ca toate celulele se divid pe acelasi plan si intr-o singura directie, ramanand apoi atasate una de alta.
Cocii care se grupeaza in acest fel formeaza in sistematica genul Streptococcus.
Lungimea lanturilor variaza de la o specie la alta;
unii streptococi se grupeaza frecvent in lanturi lungi, formate adeseori din cateva zeci de celule;
altii se grupeaza obisnuit in lanturi mai scurte, formate uneori numai din cateva celule.
Lungimea lanturilor este in parte conditionata de mediul in care au fost cultivati germenii si de natura materialului patologic.
In culturile pe medii lichide lanturile sunt de regula mult mai lungi decat in culturile dezvoltate pe suprafata mediilor semilichide (geloza cu ser)
Gruparea in tetrada
(tetra = patru).
Tetrada este o grupare de cate patru coci, dispusi intr-un aranjament regulat (sau in gramezi constituite din mai multe tetrade).
germenii se divid succesiv totdeauna pe doua planuri perpendiculare, ramanand apoi atasati in grupari de patru sau multiplu de patru.
Gruparea in octada
Se caracterizeaza prin constituirea de grupari formate din opt coci, dispusi intr-un aranjament cubic: patru intr-un prim plan si alti patru intr-un plan profund si simetric.
diviziunea celulelor se produce succesiv totdeauna pe trei planuri perpendiculare, orientate in cele trei directii ale spatiului.
Speciile care se grupeaza astfel formeaza in sistematica genul Sarcina.
Gruparea in gramezi neregulate (staphylos = ciorchine).
caracteristica stafilococilor - nse grupeaza in gramezi neregulate, mai mari sau mai mici, asemanatoare unor ciorchini de strugure
celulele se divid pe diverse planuri si directii, ramanand apoi atasate in gramezi fara un aranjament caracteristic.
La bacili se pot intalni, de asemenea, modalitati de grupare mai mult sau mai putin caracteristice cum sunt:
Diplobacilii
-sunt grupari formate din cate doi bacili dispusi cap la cap.
-doua celule rezultate din diviziune raman de regula atasate una de alta.
In cazul bacilului rujetului formatiunile diplobacilare au aspect caracteristic, germenii dispunandu-se de regula in unghi obtuz, asemanator literei V.
Streptobacilii grupari constituite din mai multi bacili dispusi cap la cap, intr-un aranjament liniar, formand astfel o insiruire de germeni in lant
Lungimea lanturilor este variabila, in functie de mediul in care germenii au fost cultivati sau de materialul patologic in care se gasesc.
Acest mod de grupare este caracteristic pentru Bacillus anthracis, care in culturile pe medii artificiale se gaseste frecvent dispus in lanturi uneori foarte lungi.
Din contra, in frotiurile facute din sangele si organele animalelor moarte de antrax, germenii se gasesc izolati sau grupati in lanturi scurte, formate din cel mult cateva elemente.
Gruparea in "grilaj" sau "palisada"
intalnita la genurile Mycobacterium si Corynebacterium
dispunerea mai multor bacili intr-un aranjament paralel, asemanator degetelor de la o manusa sau a scandurilor dintr-un gard
bacilii rezultati din diviziuni succesive sufera un proces de basculare de 180° in jurul peretelui de septare, celulele fiice luand astfel o pozitie paralela
DIMENSIUNILE
Bacteriile au dimensiuni de ordinul micronilor, cuprinse intre limite de 0,1-15 µm.
determinate genetic, constituind un important criteriu de diagnostic
De exemplu, diametrul celor mai mici coci oscileaza in jurul a 0,6 µm, iar la cocii mari in jurul a 1,2-1,5 µm (stafilococii 0,8-1µm; streptococii 0,6-1µm).
La bacili se apreciaza lungimea si grosimea
Cei mai mici bacili - Brucella (grosimea in jurul a 0,1 µm si lungimea in jurul a 0,3 µm)
- micoplasme si rickettsii - dimensiuni foarte mici (0,1-0,2 µm).
Una din cele mai mari specii bacteriene patogene este Clostridium septicum,
Spirochetele au o grosime mica, dar o lungime apreciabila (Leptospira - grosimea de 0,25-0,3 µm, lungimea variaza intre 6-20 µm)
VOLUMUL, GREUTATEA SI DENSITATEA
In raport cu dimensiunile pe care le au, volumul si greutatea sunt foarte mici
iar densitatea bacteriilor oscileaza de regula in jurul a 1,1.
De aceea ele sedimenteaza cand sunt suspendate in mediile lichide cu o densitate mai mica.
AFINITATI TINCTORIALE
Bacteriile manifesta o aviditate pronuntata pentru colorantii bazici: albastru de metilen, albastru de toluidina, cristal violet, fuxina bazica
Evidentierea acestor afinitati tinctoriale are o mare utilitate practica, in lucrarile curente de identificare a bacteriilor
Fixarea rapida, intensa si uniforma a colorantului - celulele bacteriene aparand colorate de regula omogen - investigatie optica, in celula bacteriana nu se poate evidentia prezenta unor organite citoplasmatice sau a aparatului nuclear
Afinitati tinctoriale
- comportament diferit la acelasi colorant
- intensitatii diferita fixeaza si retin colorantii utilizati
Pentru evidentierea principalelor afinitati tinctoriale ale bacteriilor sunt utilizate de regula doua metode de colorare:
- metoda Gram
- metoda Ziehl-Neelsen
Metoda de colorare Gram - doua mari categorii:
- bacterii Gram pozitive, care se coloreaza in albastru
- bacterii Gram negative care se coloreaza in rosu
Metoda de colorare Ziehl-Neelsen - pt. bacteriile acido-rezistente (M. tuberculosis, M. paratuberculosis, M. leprae etc.) doua categorii
- bacterii acido-alcoolo-rezistente, care se coloreaza in rosu
- bacterii neacido-alcoolo-rezistente, care se coloreaza in albastru
Afinitatea tinctoriala
-de natura componentelor endocelulare si mai ales de deosebirile de structura morfochimica ale peretilor celulari si ale membranei citoplasmatice
-pH ale mediului endocelular (bacteriile Gram-negative au un pH endocelular in jur de 5, in timp ce pH-ul endocelular al bacteriilor Gram-pozitive este in jur de 2 - afinitate pentru coloranti bazici = violet de gentiana)
MORFOLOGIE BACTERIANA
Bacteriile sunt vietuitoare unicelulare
Celula este protejata de un perete (membrana ) cu o structura rigida
care confera fiecarei specii o forma caracteristica.
Se disting patru tipuri morfologice principale:
Multiplicarea bacteriilor - prin sciziparitate,
aparatul nuclear se divide pe cale amitotica (prin replicarea filamentului de ADN)
* Exceptional, anumite grupe de bacterii se pot multiplica si pe alte cai (inmugurire,fragmentare etc.).
Bacteriile sunt lipsite de pigmenti fotosintetizanti
* cu exceptia catorva grupe de bacterii
Energia necesara activitatii vitale si-o procura prin reactii chimice exergonice.
vietuitoare strict chemotrofe (chemosintetizante).
poseda un bogat echipament enzimatic, care le permite sa utilizeze, in metabolismul plastic si energetic, componente organice si anorganice foarte variate.
Lumea bacteriana poate fi impartita in doua categorii:
bacterii autotrofe substrat nutritiv numai componente de natura anorganica;
bacterii heterotrofe - obligatoriu surse organice de C si N, dar pot utiliza si diferiti compusi anorganici.
Caracterele morfologice proprii unor specii sau grupe de bacterii au o
Servesc ca importante criterii taxonomice
Utilitate practica esentiala in lucrarile curente de diagnostic bacteriologic.
FORMA
Patru tipuri morfologice:
Cocii
Au o forma sferoidala, aparent rotunda, uneori usor ovalara sau chiar neregulata.
Unele specii de coci au o forma particulara caracteristica;
Streptococcus pneumoniae (pneumococ) - celulele bacteriene au forma ovalara lanceolata
Neisseria gonorrhoeae (gonococ) o forma ovalara reniforma.
Bacilii
Au forma cilindrica, mai mult sau mai putin alungita, asemanatoare unui bastonas.
La unele specii forma bacilara este usor aplatizata (Erysipelothrix insidiosa)
Numeroase specii de bacili prezinta obisnuit aspecte de polimorfism:
-forme cocoide
-forme cocobacilare,
-forme naviculare, forme bacilare foarte mult alungite care uneori au aspect filamentos etc.
Intre bacili exista deosebiri in privinta grosimii si lungimii sau a formei pe care o au capetele acestora.
La unele specii formele bacilare sunt subtiri, fine, de regula rectilinii (E. insidiosa),
la altele pot fi usor incurbate (Mycobacterium tuberculosis);
La majoritatea speciilor capetele bacililor sunt usor rotunjite
B. anthracis capetele sunt insa drepte.
La speciile din genul Fusobacterium celula se subtiaza spre capete dand bacteriilor un aspect fusiform.
Vibrionii
Au forma alungita si incurbata, uneori cu ondulatii neregulate.
Speciile din genul Vibrio au obisnuit o forma incurbata, asemanatoare cu o virgula sau cu un corn.
Spirochetele
Au aspect filiform, filamentele fiind foarte subtiri si lungi, cu numeroase ondulatii sub forma de spire regulate (asemanatoare unui tirbuson).
Aspectul spirelor poate fi diferit:
-spire numeroase (cateva zeci) foarte stranse si regulate;
-spire mai largi si mai putin regulate.
Aspectul spirelor poate fi diferit:
MOD DE GRUPARE
Celulele rezultate din diviziune se separa, raspandindu-se neuniform in mediu.
La unele bacterii celulele raman insa atasate una de alta, de o maniera care duce la formarea de grupari caracteristice
Modul de grupare constituie la unele specii un important criteriu taxonomic
Cauzele care determina constituirea de grupari caracteristice sunt multiple
Felul in care se realizeaza diviziunea celulelor bacteriene - la unele specii totdeauna pe anumite planuri si directii determinate genetic
Mentinerea lor atasata se realizeaza in majoritatea cazurilor printr-o substanta vascoasa, secretata numai pe anumite zone de la suprafata bacteriei, determinate genetic
Cele mai caracteristice modalitati de grupare sunt intalnite la coci
Diplococii (diplos = dublu). Sunt coci grupati cate doi.
fiecare celula bacteriana se divide totdeauna pe un singur plan, iar celulele fiice raman in continuare atasate, realizandu-se astfel grupari de cate doua celule sau inlantuiri scurte de asemenea diplococi.
La unele specii, aranjamentul fata in fata a celor doi coci se face de o maniera foarte caracteristica.
Str. pneumoniae (pneumococul) are o forma lanceolata si se dispune totdeauna cu partea ascutita fata in fata,
Neisseria gonorrhoeae (gonococul) are forma reniforma si se dispune totdeauna cu partea reniforma fata in fata.
Streptococii (streptos = lant) - coci dispusi intr-o inlantuire liniara, asemanatoare unui sirag de margele
Gruparea in lant este determinata de faptul ca toate celulele se divid pe acelasi plan si intr-o singura directie, ramanand apoi atasate una de alta.
Cocii care se grupeaza in acest fel formeaza in sistematica genul Streptococcus.
Lungimea lanturilor variaza de la o specie la alta;
Unii streptococi se grupeaza frecvent in lanturi lungi, formate adeseori din cateva zeci de celule;
altii se grupeaza obisnuit in lanturi mai scurte, formate uneori numai din cateva celule.
Lungimea lanturilor este in parte conditionata de mediul in care au fost cultivati germenii si de natura materialului patologic.
In culturile pe medii lichide lanturile sunt de regula mult mai lungi decat in culturile dezvoltate pe suprafata mediilor semilichide (geloza cu ser)
Gruparea in tetrada
(tetra = patru).
Tetrada este o grupare de cate patru coci, dispusi intr-un aranjament regulat (sau in gramezi constituite din mai multe tetrade).
Germenii se divid succesiv totdeauna pe doua planuri perpendiculare, ramanand apoi atasati in grupari de patru sau multiplu de patru.
Gruparea in octada
Se caracterizeaza prin constituirea de grupari formate din opt coci, dispusi intr-un aranjament cubic: patru intr-un prim plan si alti patru intr-un plan profund si simetric.
diviziunea celulelor se produce succesiv totdeauna pe trei planuri perpendiculare, orientate in cele trei directii ale spatiului.
Speciile care se grupeaza astfel formeaza in sistematica genul Sarcina.
Gruparea in gramezi neregulate (staphylos = ciorchine).
caracteristica stafilococilor - nse grupeaza in gramezi neregulate, mai mari sau mai mici, asemanatoare unor ciorchini de strugure
celulele se divid pe diverse planuri si directii, ramanand apoi atasate in gramezi fara un aranjament caracteristic.
La bacili se pot intalni, de asemenea, modalitati de grupare mai mult sau mai putin caracteristice cum sunt:
Diplobacilii
sunt grupari formate din cate doi bacili dispusi cap la cap.
doua celule rezultate din diviziune raman de regula atasate una de alta.
In cazul bacilului rujetului formatiunile diplobacilare au aspect caracteristic, germenii dispunandu-se de regula in unghi obtuz, asemanator literei V.
Streptobacilii
Grupari constituite din mai multi bacili dispusi cap la cap, intr-un aranjament liniar, formand astfel o insiruire de germeni in lant.
Lungimea lanturilor este variabila, in functie de mediul in care germenii au fost cultivati sau de materialul patologic in care se gasesc.
Acest mod de grupare este caracteristic pentru Bacillus anthracis, care in culturile pe medii artificiale se gaseste frecvent dispus in lanturi uneori foarte lungi.
Din contra, in frotiurile facute din sangele si organele animalelor moarte de antrax, germenii se gasesc izolati sau grupati in lanturi scurte, formate din cel mult cateva elemente.
Gruparea in "grilaj" sau "palisada"
Intalnita la genurile Mycobacterium si Corynebacterium dispunerea mai multor bacili intr-un aranjament paralel, asemanator degetelor de la o manusa sau a scandurilor dintr-un gard.
Bacilii rezultati din diviziuni succesive sufera un proces de basculare de 180° in jurul peretelui de septare, celulele fiice luand astfel o pozitie paralela
DIMENSIUNILE
Bacteriile au dimensiuni de ordinul micronilor, cuprinse intre limite de 0,1-15 µm.
determinate genetic, constituind un important criteriu de diagnostic
De exemplu, diametrul celor mai mici coci oscileaza in jurul a 0,6 µm, iar la cocii mari in jurul a 1,2-1,5 µm (stafilococii 0,8-1µm; streptococii 0,6-1µm).
La bacili se apreciaza lungimea si grosimea
Cei mai mici bacili - Brucella (grosimea in jurul a 0,1 µm si lungimea in jurul a 0,3 µm)
- micoplasme si rickettsii - dimensiuni foarte mici (0,1-0,2 µm).
Una din cele mai mari specii bacteriene patogene este Clostridium septicum,
Spirochetele au o grosime mica, dar o lungime apreciabila (Leptospira - grosimea de 0,25-0,3 µm, lungimea variaza intre 6-20 µm)
VOLUMUL, GREUTATEA SI DENSITATEA
In raport cu dimensiunile pe care le au, volumul si greutatea sunt foarte mici
iar densitatea bacteriilor oscileaza de regula in jurul a 1,1.
De aceea ele sedimenteaza cand sunt suspendate in mediile lichide cu o densitate mai mica.
AFINITATI TINCTORIALE
Bacteriile manifesta o aviditate pronuntata pentru colorantii bazici: albastru de metilen, albastru de toluidina, cristal violet, fuxina bazica
Evidentierea acestor afinitati tinctoriale are o mare utilitate practica, in lucrarile curente de identificare a bacteriilor.
Fixarea rapida, intensa si uniforma a colorantului - celulele bacteriene aparand colorate de regula omogen - investigatie optica, in celula bacteriana nu se poate evidentia prezenta unor organite citoplasmatice sau a aparatului nuclear
Afinitati tinctoriale
- comportament diferit la acelasi colorant
- intensitatii diferita fixeaza si retin colorantii utilizati
Pentru evidentierea principalelor afinitati tinctoriale ale bacteriilor sunt utilizate de regula doua metode de colorare:
- metoda Gram
- metoda Ziehl-Neelsen
Metoda de colorare Gram - doua mari categorii:
- bacterii Gram pozitive, care se coloreaza in albastru
- bacterii Gram negative care se coloreaza in rosu
Metoda de colorare Ziehl-Neelsen - pt. bacteriile acido-rezistente (M. tuberculosis, M. paratuberculosis, M. leprae etc.) doua categorii
- bacterii acido-alcoolo-rezistente, care se coloreaza in rosu
- bacterii neacido-alcoolo-rezistente, care se coloreaza in albastru
Afinitatea tinctoriala
-de natura componentelor endocelulare si mai ales de deosebirile de structura morfochimica ale peretilor celulari si ale membranei citoplasmatice
-pH ale mediului endocelular (bacteriile Gram-negative au un pH endocelular in jur de 5, in timp ce pH-ul endocelular al bacteriilor Gram-pozitive este in jur de 2 - afinitate pentru coloranti bazici = violet de gentiana)
STRUCTURA CELULEI BACTERIENE
Morfostructura celulei bacteriene se disting:
Organitele celulare prezente numai la unele specii bacteriene:
pili,
cili,
capsula
PERETELE CELULEI BACTERIENE (MEMBRANA
O formatiune anatomica bine individualizata morfologic si chimic, relativ compacta, rigida si foarte rezistenta ce protejeaza la exterior celula bacteriana
De organizarea si natura componentelor parietale depinde in mare masura comportamentul bacteriilor fata de factorii fizico-chimici si biologici.
Structura morfo-chimica a peretelui celulei bacteriene
o formatiune anatomica bine cunoscuta morfo-chimic, relativ compacta, rezistenta si rigida
Grosimea - 15-35 nm, fiind de regula mai subtire la celulele tinere.
Are o grosime mai mare la bacteriile Gram-pozitive decat la bacteriile Gram-negative.
Cantitativ reprezinta la unele specii pana la 25% din volumul celulei.
electrono-optic - structura fundamentala se aseamana cu o plasa sau retea, cu trama si ochiuri regulate.
Membrana bacteriana apare alcatuita din fascicule cu fibre paralele care se incruciseaza, formand in jurul celulei o veritabila retea continua.
Are calitatea de membrana semipermeabila, cu structura poroasa, care permite realizarea schimburilor active de metaboliti celula si mediu, in ambele sensuri.
La toate bacteriile, in structura peretelui intra un component macromolecular denumit mureina (sau mucocomplex, peptidoglican, muco-peptid etc.)
O configuratie spatiala in "retea" - bi sau tridimensionala
Formeaza in jurul celulei un veritabil saccul mureinic, a carui rezistenta si rigiditate sunt consolidate prin legaturi covalente cu alte componente parietale.
Din punct de vedere chimic mureina este formata dintr-un polimer glucidic liniar, rezultat din condensarea alternativa a acetil-glucozaminei si a acidului acetil-muramic.
Aceasta formeaza scheletul macromoleculei, rezistenta sa fiind data de legaturile glicozidice dintre cele doua componente.
La acidul acetil-muramic se leaga lateral oligopeptide (tetra sau pentapeptide)
Intre oligopeptidele invecinate se stabilesc legaturi chimice aditionale (covalente), care consolideaza rezistenta macromoleculei.
Pe langa mureina, peretele contine numeroase alte componente, de regula proprii fiecarei specii bacteriene
Prin dispunerea acestor componente in perete, ele formeaza in principiu doua straturi:
- stratul R (rigid), alcatuit predominant din mureina, ocupa zona profunda a peretelui
- stratul P "plastic" format din diverse componente glucidice, lipidice si proteice etc., specifice fiecarei bacterii
In privinta structurii peretelui exista insa mari deosebiri in special de la un grup de bacterii la altul
Peretele bacteriilor Gram pozitive
Polizaharidele caracteristice pentru bacteriile Gram-pozitive sunt:
acizii teichoici
Acizii teichoici (gr. teichos = zid)
-sunt polimeri liniari de glicerol-fosfat sau ribitol-fosfat, lungi si flexibili.
-numarul subunitatilor constitutive difera in functie de natura lor si de specia bacteriana.
-se gasesc numai la bacteriile Gram-pozitive, in cantitati relativ mari (40-60% din greutatea uscata a peretelui)
-la gruparile hidroxid ale acestor polimeri liniari se leaga alte componente chimice (glucoza, lactoza, N-acetilglucozamina etc.) proprii fiecarei specii.
Asemenea componente indeplinesc rol in determinarea specificitatii antigenice.
Se gasesc situati in perete mai mult sau mai putin profund.
In raport cu pozitia si natura lor, se disting trei categorii:
acizii teichoici parietali
acizii teichuronici
teichoici membranari (lipoteichoici)
acizii teichoici parietali
sunt componente de suprafata
- legate de regula covalent la mureina,
contribuie la consolidarea structurii si rezistentei acesteia si a peretelui
Variaza cantitativ in functie de conditiile de cultivare si de mediu
La gruparile hidroxid ale acestora se leaga diferite alte componente, formand impreuna antigenii de suprafata;
acizii teichuronici
Formati din unitati dizaharidice repetate, la care se leaga covalent N-acetil-glucozamina si acidul glucuronic.
Se gasesc in perete asociati cu acizii teichoici.
acizii teichoici membranari (lipoteichoici)
Sunt formati prin polimerizarea a 20-30 unitati de glicerol-fosfat.
Se gasesc situati in zona profunda, in aria "dintre perete si membrana citoplasmatica, unde formeaza o retea.
Sunt de regula legati covalent de componentele membranei citoplasmatice.
Cand peretele este inlaturat, ei raman ancorati la membrana citoplasmatica.
Sunt mai caracteristici pentru bacteriile Gram-pozitive decat acizii teichoici, sinteza lor nefiind influentata de conditiile de cultivare.
Rolul acizilor teichoici este deosebit de complex:
Au actiune antifagocitara (la Streptococcus pyogenes) si pot servi ca receptori pentru fagi si colicine (ac. lipoteichoici).
Peretele bacteriilor Gram negative
Stratul profund
Stratul plastic sau membrana externa
Stratul intern
Stratul profund
electronodens
-o grosime de 1,5-3,0 nm
-constituit mureina si lipoproteine
Stratul plastic sau membrana externa
Glucidele din complex lipopolizaharidic sunt orientate spre suprafata, ele fiind purtatoare ale specificitatii antigenice.
Stratul intern
Constituit din fosfolipide care se leaga prin lipoproteine la stratul mureinic profund.
Rol biologic
Prin structura sa compacta, densa si relativ rigida, confera bacteriilor forma caracteristica
Rezistenta si elasticitatea sa sunt esentiale pentru mentinerea integritatii celulei, asigurand protectia acesteia fata de socurile din mediu.
Contracareaza presiunea osmotica endocelulara, care atinge obisnuit valori medii echivalente a 15-20 atmosfere (pana la 20-30 atmosfere la cocii Gram-pozitivi; 4-8 atmosfere la bacteriile Gram-negative).
Se opune si limiteaza mecanic patrunderea in celula a unor cantitati mari de apa, puternic atrasa de hidrofilia materiei citoplasmatice (foarte bogata in saruri minerale si in alte componente hidrosolubile).
Structura poroasa confera peretelui celular caracterul de membrana semipermeabila, care permite realizarea unor schimburi continui si selective de metaboliti, intre celula si mediu.
Desi nu are rol metabolic, se admite totusi ca poate interveni in unele procese de fixare si transport ionic.
De anumite caracteristici structurale si chimice ale peretelui bacterian depind in masura considerabila afinitatile tinctoriale ale bacteriilor.
Peretele celulei bacteriene participa in procesele de diviziune celulara, creste centripet si separa complet cele doua celule fiice rezultate din diviziune
Componentele macromoleculare parietale sunt in cele mai multe cazuri antigenice, specificitatea antigenica fiind conferita de regula de componentele glucidice. De exemplu, lipopolizaharidele reprezinta antigenii "O" ai bacteriilor Gram-negative.
Componentele antigenice parietale diferentiaza si individualizeaza fiecare specie bacteriana, constituind frecvent un important criteriu taxonomic, de mare utilitate in identificarea speciilor si tipurilor bacteriene.
Unele componente parietale au rol in agresivitatea germenilor, fiind considerate factori de patogenitate. De exemplu, complexele lipopolizaharidice au rol de endotoxine - antigenii "Vi"
Prin natura sa chimica, peretele bacterian joaca un rol important in comportamentul unor bacterii fata de factorii de aparare ai organismului.
Astfel, bacilul tuberculozei rezista in citoplasma fagocitelor datorita mai ales bogatiei peretelui celular in ceride si fosfolipide.
Sensibilitatea crescuta la lizozima a bacteriilor Gram-pozitive si la alexina a bacteriilor Gram-negative este determinata, de asemenea, de caracteristicile morfo-chimice ale peretelui celular.
Anumite grupari chimice de suprafata (de exemplu, lipopolizaharidele bacteriilor Gram-negative) au rol de receptori pentru bacteriofagi. Acest fapt este atestat si de constatarea ca bacteriofagii nu se pot fixa pe suprafata protoplastilor.
Peretele bacterian determina comportamentul diferit al bacteriilor la numerosi agenti fizico-chimici si biologici. De exemplu, bacteriile Gram-pozitive sunt sensibile la penicilina si mai sensibile la acizi, detergenti, iod etc., decat bacteriile Gram-negative, dar sunt mai rezistente la alcali, agenti oxidanti, enzime proteolitice etc.
SPATIUL PERIPLASMATIC
Este caracteristic numai la bacteriile Gram-negative. Cuprinde zona profunda delimitata spre exterior de membrana externa a peretelui, iar spre interior de membrana citoplasmatica .
Prin urmare, in spatiul periplasmatic este cuprins si stratul "R" al peretelui, format din mureina si lipoproteine.
In aceasta zona se gasesc numeroase enzime (fosfataza alcalina, acid hexozofosfatazic, fosfodiesteraza ciclica, diferite hidrolaze, fosfataze, nucleaze, penicilinaze etc.), precum si anumite proteine neenzimatice, care au rol de a lega specific anumite componente (glucide, aminoacizi, ioni organici etc.) si de a le transporta la membrana citoplasmatica.
PROTOPLASTUL
Prin indepartarea peretelui bacterian se obtine o formatiune bine individualizata numita protoplast.
Acesta contine toate componentele endocelulare (citoplasma, organite citoplasmatice si aparat nuclear), delimitate la exterior de membrana citoplasmatica.
Masa protoplastilor reprezinta aproximativ 60-80% din greutatea uscata a celulei bacteriene.
Protoplasti se comporta diferit in functiei de caracteristicile chimice ale mediului.
In mediile izotone (prin adaos de zaharoza in concentratii de 0,1-0,5 M) protoplastii isi conserva integritatea morfologica, luand insa o forma izodiametrica, sferoidala, datorita fortelor osmotice endocelulare si a elasticitatii membranei citoplasmatice.
In mediile hipertonice, datorita pierderii de apa endocelulara, volumul protoplastilor se retracta, se micsoreaza.
In mediile hipotonice (cum sunt cele naturale) protoplastii se dezintegreaza imediat, intrucat absorbtia rapida de apa duce la cresterea considerabila a volumului si a presiunii osmotice endocelulare, ceea ce provoaca ruperea membranei. Se produce astfel o adevarata "explozie osmotica", continutul celular dispersandu-se in mediu.
Protoplastii pot fi obtinuti mai usor din bacteriile Gram-pozitive, al caror perete este predominant glucidic.
In acest scop se poate recurge la tratarea bacteriilor cu lizozima, care hidrolizeaza mureina si indeparteaza peretele celular.
Formatiuni asemanatoare se pot obtine si prin cultivarea bacteriilor in prezenta penicilinei, antibioticul interferand sinteza mureinei si implicit al peretelui celular. In prezenta antibioticului, celulele fiice rezultate din diviziune nu-si mai pot sintetiza integral peretele, dand astfel nastere la sferoplasti, formatiuni care mai pastreaza resturi de perete.
Datorita fragmentelor de perete ramase pe suprafata lor, sferoplastii se pot multiplica si isi pot resintetiza peretele, procesul pornind chiar de la fragmentele ramase, care indeplinesc probabil rolul de starter (de matrita pentru resintetizarea sacului mureinic).
Prin aceasta sferoplastii se deosebesc de protoplasti, care nu-si pot resintetiza peretele.
Prin ansamblul caracteristicilor sale, protoplastul reprezinta unitatea morfo-biologica de baza a celulei bacteriene, conservand toate atributele esentiale ale acesteia.
Datorita conservarii acestor atribute, protoplastii pot fi considerati celule bacteriene lipsite de perete.
Morfostructura celulei bacteriene :
PERETELE CELULEI BACTERIENE
O formatiune anatomica bine individualizata morfologic si chimic, relativ compacta, rigida si foarte rezistenta ce protejeaza la exterior celula bacteriana
De organizarea si natura componentelor parietale depinde in mare masura comportamentul bacteriilor fata de factorii fizico-chimici si biologici.
Structura morfo-chimica a peretelui celulei bacteriene
Grosimea - 15-35 nm, fiind de regula mai subtire la celulele tinere, grosime mai mare la bacteriile Gram-pozitive decat la bacteriile Gram-negative.
Cantitativ reprezinta la unele specii pana la 25% din volumul celulei.
Electrono-optic - structura fundamentala se aseamana cu o plasa sau retea, cu trama si ochiuri regulate
Membrana bacteriana apare alcatuita din fascicule cu fibre paralele care se incruciseaza, formand in jurul celulei o veritabila retea continua.
Are calitatea de membrana semipermeabila, cu structura poroasa, care permite realizarea schimburilor active de metaboliti celula si mediu, in ambele sensuri.
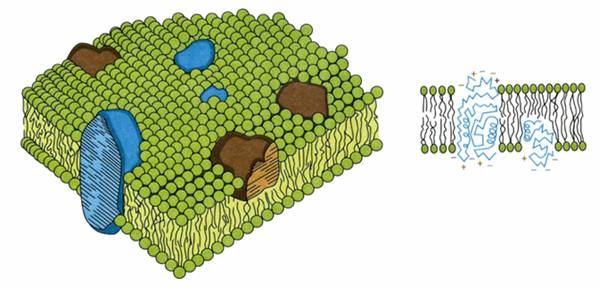
La toate bacteriile, in structura peretelui intra un component macromolecular denumit mureina (sau mucocomplex, peptidoglican, muco-peptid etc.).
O configuratie spatiala in "retea" - bi sau tridimensionala.
Formeaza in jurul celulei un veritabil saccul mureinic, a carui rezistenta si rigiditate sunt Consolidate prin legaturi covalente cu alte componente parietale.
Din punct de vedere chimic mureina este formata dintr-un polimer glucidic liniar, rezultat din condensarea alternativa a acetil-glucozaminei si a acidului acetil-muramic.
Aceasta formeaza scheletul macromoleculei, rezistenta sa fiind data de legaturile glicozidice dintre cele doua componente.
La acidul acetil-muramic se leaga lateral oligopeptide (tetra sau pentapeptide).
Intre oligopeptidele invecinate se stabilesc legaturi chimice aditionale (covalente), care consolideaza rezistenta macromoleculei.
Pe langa mureina, peretele contine numeroase alte componente, de regula proprii fiecarei specii bacteriene.
Prin dispunerea acestor componente in perete, ele formeaza in principiu doua straturi:
- stratul R (rigid), alcatuit predominant din mureina, ocupa zona profunda a peretelui
- stratul P "plastic" format din diverse componente glucidice, lipidice si proteice etc., specifice fiecarei bacterii
In privinta structurii peretelui exista insa mari deosebiri in special de la un grup de bacterii la altul
Peretele-bacteriilor
Gram pozitive
Polizaharidele caracteristice pentru bacteriile Gram-pozitive sunt:
acizii teichoici
Acizii teichoici (gr. teichos = zid)
-sunt polimeri liniari de glicerol-fosfat sau ribitol-fosfat, lungi si flexibili
-numarul subunitatilor constitutive difera in functie de natura lor si de specia bacteriana
-se gasesc numai la bacteriile Gram-pozitive, in cantitati relativ mari (40-60% din greutatea uscata a peretelui)
-la gruparile hidroxid ale acestor polimeri liniari se leaga alte componente chimice (glucoza, lactoza, N-acetilglucozamina etc.) proprii fiecarei specii.
Asemenea componente indeplinesc rol in determinarea specificitatii antigenice.
Se gasesc situati in perete mai mult sau mai putin profund.
In raport cu pozitia si natura lor, se disting trei categorii:
acizii teichoici parietali
acizii teichuronici
teichoici membranari (lipoteichoici)
acizii teichoici parietali
sunt componente de suprafata
- legate de regula covalent la mureina, contribuie la consolidarea structurii si rezistentei acesteia si a peretelui.
Variaza cantitativ in functie de conditiile de cultivare si de mediu.
la gruparile hidroxid ale acestora se leaga diferite alte componente, formand impreuna antigenii de suprafata;
acizii teichuronici
Formati din unitati dizaharidice repetate, la care se leaga covalent N-acetil-glucozamina si acidul glucuronic.
Se gasesc in perete asociati cu acizii teichoici.
Rolul acizilor teichoici este deosebit de complex:
-confera bacteriilor Gram-pozitive o serie de proprietati
-protejeaza mureina fata de anumite enzime si detergenti.
-sunt implicati apoi in mentinerea, in zona de la suprafata membranei citoplasmatice, a unei anumite concentratii de ioni metalici
-esentiala pentru activitatea unor sisteme enzimatice membranare
-au actiune antifagocitara (la Streptococcus pyogenes) si pot servi ca receptori pentru fagi si colicine (ac. lipoteichoici).
Peretele bacteriilor Gram negative
Stratul profund
electronodens
o grosime de 1,5-3,0 nm
constituit mureina si lipoproteine
Stratul plastic sau membrana externa
o grosime de 6-20 nm
alcatuit din lipopolizaharide (50%), fosfolipide (35%) proteine (15%)
Glucidele din complex lipopolizaharidic sunt orientate spre suprafata, ele fiind purtatoare ale specificitatii antigenice
Stratul intern
constituit din fosfolipide care se leaga prin lipoproteine la stratul mureinic profund
Rol biologic al peretelui bacterian
Prin structura sa compacta, densa si relativ rigida, confera bacteriilor forma caracteristica
Rezistenta si elasticitatea sa sunt esentiale pentru mentinerea integritatii celulei, asigurand protectia acesteia fata de socurile din mediu.
Contracareaza presiunea osmotica endocelulara, care atinge obisnuit valori medii echivalente a 15-20 atmosfere (pana la 20-30 atmosfere la cocii Gram-pozitivi; 4-8 atmosfere la bacteriile Gram-negative).
Se opune si limiteaza mecanic patrunderea in celula a unor cantitati mari de apa, puternic atrasa de hidrofilia materiei citoplasmatice (foarte bogata in saruri minerale si in alte componente hidrosolubile).
Structura poroasa confera peretelui celular caracterul de membrana semipermeabila, care permite realizarea unor schimburi continui si selective de metaboliti, intre celula si mediu. Desi nu are rol metabolic, se admite totusi ca poate interveni in unele procese de fixare si transport ionic.
De anumite caracteristici structurale si chimice ale peretelui bacterian depind in masura considerabila afinitatile tinctoriale ale bacteriilor.
Peretele celulei bacteriene participa in procesele de diviziune celulara, creste centripet si separa complet cele doua celule fiice rezultate din diviziune.
Componentele macromoleculare parietale sunt in cele mai multe cazuri antigenice, specificitatea antigenica fiind conferita de regula de componentele glucidice.
De exemplu, lipopolizaharidele reprezinta antigenii "O" ai bacteriilor Gram-negative.
Componentele antigenice parietale diferentiaza si individualizeaza fiecare specie bacteriana, constituind frecvent un important criteriu taxonomic, de mare utilitate in identificarea speciilor si tipurilor bacteriene.
Unele componente parietale au rol in agresivitatea germenilor, fiind considerate factori de patogenitate. De exemplu, complexele lipopolizaharidice au rol de endotoxine, iar antigenii "Vi" pe care ii au unele salmonele favorizeaza virulenta
Prin natura sa chimica, peretele bacterian joaca un rol important in comportamentul unor bacterii fata de factorii de aparare ai organismului. Astfel, bacilul tuberculozei rezista in citoplasma fagocitelor datorita mai ales bogatiei peretelui celular in ceride si fosfolipide. Sensibilitatea crescuta la lizozima a bacteriilor Gram-pozitive si la alexina a bacteriilor Gram-negative este determinata, de asemenea, de caracteristicile morfo-chimice ale peretelui celular.
Anumite grupari chimice de suprafata (de exemplu, lipopolizaharidele bacteriilor Gram-negative) au rol de receptori pentru bacteriofagi.
Peretele bacterian determina comportamentul diferit al bacteriilor la numerosi agenti fizico-chimici si biologici.
De exemplu, bacteriile Gram-pozitive sunt mai sensibile la acizi, detergenti, iod etc., decat bacteriile Gram-negative, dar sunt mai rezistente la alcali, agenti oxidanti, enzime proteolitice etc.
SPATIUL PERIPLASMATIC
Este caracteristic numai la bacteriile Gram-negative
Cuprinde zona profunda delimitata spre exterior de membrana externa a peretelui, iar spre interior de membrana citoplasmatica .
Prin urmare, in spatiul periplasmatic este cuprins si stratul "R" al peretelui, format din mureina si lipoproteine.
In aceasta zona se gasesc
-numeroase enzime (fosfataza alcalina, acid hexozofosfatazic, fosfodiesteraza ciclica, diferite hidrolaze, fosfataze, nucleaze, penicilinaze etc.)
-proteine neenzimatice, care au rol de a lega specific anumite componente (glucide, aminoacizi, ioni organici etc.) si de a le transporta la membrana citoplasmatica.
PROTOPLASTUL
O formatiune bine individualizata care se obtine prin indepartarea peretelui bacterian.
Acesta contine toate componentele endocelulare (citoplasma, organite citoplasmatice si aparat nuclear), delimitate la exterior de membrana citoplasmatica.
Masa protoplastilor reprezinta aproximativ 60-80% din greutatea uscata a celulei bacteriene.
Protoplasti se comporta diferit in functiei de caracteristicile chimice ale mediului.
In mediile izotone (prin adaos de zaharoza in concentratii de 0,1-0,5 M) protoplastii isi conserva integritatea morfologica, luand insa o forma izodiametrica, sferoidala, datorita fortelor osmotice endocelulare si a elasticitatii membranei citoplasmatice.
In mediile hipertone, datorita pierderii de apa endocelulara, volumul protoplastilor se retracta, se micsoreaz
In mediile hipotonice (cum sunt cele naturale) protoplastii se dezintegreaza imediat, intrucat absorbtia rapida de apa duce la cresterea considerabila a volumului si a presiunii osmotice endocelulare, ceea ce provoaca ruperea membranei.
Se produce astfel o adevarata "explozie osmotica", continutul celular dispersandu-se in mediu.
Protoplastii pot fi obtinuti mai usor din bacteriile Gram-pozitive, al caror perete este predominant glucidic.
In acest scop se poate recurge la tratarea bacteriilor cu lizozima, care hidrolizeaza mureina si indeparteaza peretele celular.
Formatiuni asemanatoare se pot obtine si prin cultivarea bacteriilor in prezenta penicilinei, antibioticul interferand sinteza mureinei si implicit al peretelui celular. In prezenta antibioticului, celulele fiice rezultate din diviziune nu-si mai pot sintetiza integral peretele, dand astfel nastere la sferoplasti, formatiuni care mai pastreaza resturi de perete.
Datorita fragmentelor de perete ramase pe suprafata lor, sferoplastii se pot multiplica si isi pot resintetiza peretele, procesul pornind chiar de la fragmentele ramase, care indeplinesc probabil rolul de starter (de matrita pentru resintetizarea sacului mureinic).
Prin aceasta sferoplastii se deosebesc de protoplasti, care nu-si pot resintetiza peretele.
Prin ansamblul caracteristicilor sale, protoplastul reprezinta unitatea morfo-biologica de baza a celulei bacteriene, conservand toate atributele esentiale ale acesteia.
Datorita conservarii acestor atribute, protoplastii pot fi considerati celule bacteriene lipsite de perete.
Morfostructura celulei bacteriene
-ribozom
-vacuole de apa
-granule sau corpusculi metacromatici
Organitele celulare prezente numai la unele specii bacteriene:
- pili,
- cili,
- capsula
MEMBRANA CITOPLASMATICA
Poate fi pusa in evidenta numai prin mijloace speciale de investigatii.
Se muleaza perfect la fata interna a peretelui, cu care are doar raporturi de contiguitate.
In schimb, raporturile sunt foarte stranse cu masa citoplasmatica.
Structura morfo-chimica
Electrono-optic
-o formatiune triplu stratificata
-doua straturi electrono-opace,separate de un strat median electrono-transparent.
Matricea membranei citoplasmatice este formata din fosfolipide dispuse in strat dublu, moleculele lor avand o structura complexa, cu o extremitate polara hidrofila si alta hidrofoba.
-dispunerea - maniera caracteristica:
-cu extremitatile polare hidrofobe orientate spre interior, respectiv fata in fata, acestea formand (stratul median electrono-transparent);
-extremitatile ionice hidrofile, orientate spre exterior, formeaza cele doua straturi electrono-opace
O asemenea aranjare confera membranei citoplasmatice rolul de bariera osmotica si de filtru selectiv, ea fiind penetrabila numai la moleculele liposolubile si nepenetrabila la moleculele hidrosolubile.
In stratul fluid dublu lipidic plutesc moleculele proteice globulare
ele se pot deplasa liber, fiind orientate cu regiunile hidrofile spre suprafata.
De unele proteine sunt legate cantitati mici de glucide (polizaharide).
Aceste proteine membranare sunt de doua tipuri:
integrate (trans-membranare) care strabat toata grosimea membranei
de suprafata (periferice), situate in stratul lipidic extern sau in cel intern, de regula legate de proteinele integrate.
Rolul lor este multiplu:
Rolul biologic al membranei citoplasmatice
Bariera osmotica - structura morfo-chimica ii confera proprietati de permeabilitate selectiva.
La nivelul ei activeaza numeroase enzime de tipul permeazelor, cu rol in fixarea si transportul metabolitilor in ambele sensuri.
Membrana citoplasmatica permite iesirea din celula a unor componente proteice, cum sunt exotoxinele si exoenzimele, intervenind totodata in reglarea acestor procese.
Faptul este confirmat de constatarea ca, membrana citoplasmatica poate retine in celula unele exoenzime, proces definit ca fenomen de cripticitate.
Rol in metabolismul energetic.
La nivelul membranei citoplasmatice sunt localizate enzime respiratorii numeroase de tipul citocromilor;
Este sediul unor procese de sinteza
In membrana se realizeaza sinteza unor componente, care intra in structura peretelui bacterian, a capsulei, a cililor etc. ;
In cursul procesului de diviziune participa la sinteza septumului care separa continutul endocelular.
CITOPLASMA
Reprezinta cantitativ componentul major al celulei bacteriene.
Raportul dintre aceasta si materia nucleara variaza intre 2:1-10:1, in functie de specie si de starea fiziologica a celulei.
Caracteristici morfo-structurale
Caracteristici fizico-chimice
Caracteristici morfo-structurale:
Omogenitatea si densitatea materiei citoplasmatice variaza:
Caracteristici fizico-chimice
Materia citoplasmatica are un pronuntat caracter hidric
Un sistem coloidal complex - se gasesc in stare de dispersie coloidala numeroase componente moleculare organice si anorganice (proteine, glucide, lipide si saruri minerale).
Foarte bogata in special in proteine (foarte multe enzime) si in ribonucleoproteine - reactie pronuntata acida - pH = 2 - 3.
Mezozomii
Sunt structuri membranare intracitoplasmatice, cu aspect de sac sau punga, formate prin cresterea si invaginarea membranei citoplasmatice.
Dupa localizarea lor se disting trei tipuri de mezozomi:
septali - cu septul de diviziune;
nucleari - cu cromozomul celulei;
periferici - cu formarea sporului.
La bacteriile Gram-pozitive - mai numerosi si mai bine dezvoltati.
La bacteriile Gram-negative - in numar mai mic,rudimentari, slab dezvoltati, mai greu de observat.
Aspectul morfologic disting trei tipuri:
mezozomi lamelari - formati prin cresterea, invaginarea si plierea membranei citoplasmatice in spirale rasucite ca un ghem;
mezozomi tubulari - constituiti din formatiuni tubulare, mai mult sau mai putin lungi si incolacite;
mezozomi veziculari (saciformi) - cu aspect de vezicule aproape sferice.
Mezozomii au numeroase functii:
-maresc considerabil suprafata membranei citoplasmatice, ceea ce influenteaza favorabil activitatea celulara, ca raspuns la conditiile oferite de mediu.
-in replicarea si segregarea cromozomului si in transmiterea de la suprafata celulei a semnalului biochimic care initiaza replicarea.
-sunt organite echivalente mitocondriilor, la nivelul mezozomilor fiind concentrate cantitati mari de enzime respiratorii, implicate in procese de:
Ribosomii
Organite intracitoplasmatice de forma aproximativ sferica, rol esential in metabolismul proteic, la nivelul lor realizandu-se sinteza proteinelor bacteriene.
Din punct de vedere chimic sunt complexe macromoleculare ribo-nucleo-proteinice, formate din 55 molecule proteinice si 3 molecule de ARN.
Se apreciaza ca o celula poate contine intre 1.500-100.000 de ribosomi - procesele de biosinteza a proteinelor in celula bacteriana.
Majoritatea ribosomilor plutesc liber in citoplasma, dar o parte se gasesc plasati la fata interna a membranei citoplasmatice.
O mare parte sunt legati in formatiuni poliribosomale, la nivelul carora se realizeaza sinteza polipeptidelor, ceea ce implica traducerea informatiei genetice adusa de ARNm si translatia acesteia in structura polipeptidei, respectiv in ordinea de succesiune a acizilor aminati.
In citoplasma unor bacterii pot exista vacuole de apa si granule sau corpusculi metacromatici
Vacuolele, sunt izolate de materia citoplasmatica printr-o semimembrana numita tonoplast (formata prin condensarea de proteine si lipide).
dimensiunile - intre 0,3-0,6 µm.
Rol:
-rezerve endocelulare de apa - celulele in plina activitate metabolica, isi autoregleaza prompt echilibrul ionic
-osmotic - la solvarea si acumularea in vacuola a unor metaboliti sau toxice
Granulatiile si corpusculii metacromatici
Depozite de substante de rezerva (glucide, lipide, metafosfati), sub forma de polimeri insolubili in mediul hidric endocelular, cu aspect granular sau corpuscular.
Polimerizarea se realizeaza prin legaturi fosfoergice in care se inmagazineaza cantitati mari de energie.
Asemenea polimeri servesc ca surse de C, P si alte elemente, ca surse de energie, fiecare etapa a depolimerizarii eliberand cantitati apreciabile de energie calorica.
- Granulele glucidice
- Granulele lipidice
Granulele de metafosfati sau de volutina
Granulele glucidice
sub forma de granule de amidon sau de glicogen (bacteriile coliforme) granulatii fine, uneori vizibile numai prin microscopie electronica identificarea lor se poate face biochimic ( prin colorare cu iod, granulele de glicogen colorandu-se spre brun, cele de amidon in albastru, iar citoplasma bacteriana in galben)
Granulele lipidice
polimerizarea acidului betahidroxibutiric
au dimensiuni mai mari, vizibile la microscopul optic, mai ales la microscopul cu contrast de faza, evidentiere prin colorarea selectiva cu negru de Sudan.
Granulele de metafosfati sau de volutina
prin polimerizarea metafosfatului, care impreuna cu ARN dau complexe macromoleculare insolubile.
vizibile la microscopul optic - forma unor corpusculi dimensiuni mari, pana la 0,6 µm
au afinitati pentru colorantii bazici albastri (albastru de toluidina, albastru de metilen) manifestand proprietati de metacromazie (corpusculii se coloreaza in rosu, iar soma bacteriana in albastru).
NUCLEUL
nu poate fi pus in evidenta prin metodele uzuale de examinare si colorare
nucleul bacterian este mai putin individualizat si evoluat, lipsit de
membrana nucleara
de o structura interna propriu-zisa.
definirea sa prin termenii de: nucleoid, nucleozom, genom, cromozom etc.
formatiunea nucleara se gaseste ancorata la un mezozom din zona ecuatoriala a celulei, iar prin intermediul acestuia la membrana citoplasmatica
nucleul bacterian este format de fapt dintr-un singur filament foarte lung de ADN, dublu helicoidal, care formeaza o macromolecula giganta
Capetele filamentului sunt legate unul de altul, dand astfel macromoleculei de ADN o configuratie circulara, aceasta reprezentand si cromozomul unic, circular, al celulei bacteriene.
Ca urmare, avand un singur cromozom, bacteriile sunt definite ca vietuitoare cu organizare genetica haploida.
Multiplicarea nucleului se face pe cale amitotica, prin replicarea (duplicarea) ADN-ului.
Procesul consta in scindarea macromoleculei de ADN in cele doua catene, fiecare catena servind apoi ca matrita la sinteza complementara a catenei congenere
In final rezulta doua macromolecule identice de ADN, in care se conserva nealterata intreaga informatie genetica detinuta in cromozom de celula mama.
STRUCTURI EXTRAPARIETALE
glicocalixul
invelisul mucos
capsula
sintetizate in anumite conditii de mediu
lipsa lor nu afecteaza viabilitatea celulei bacteriene
prezenta lor confera bacteriilor anumite caracteristici si avantaje biologice
Glicocalixul
retea de fibre polizaharidice, asemanatoare cu o pasla, care ajuta bacteria sa se atasee la suprafata unor substraturi pe care se dezvolta si le colonizeaza
bacteriile patogene - traiesc si se multiplica pe suprafata mucoaselor intestinale, a cailor urinare, in cavitatea bucala
Invelisul mucos
un invelis amorf constituit din substante insolubile sau putin solubile, sintetizate de unele bacterii, in medii lichide, care se depun la suprafata peretelui celular sub forma unei mase neorganizate de materie mucoasa
la cele mai multe specii - subtire, fin, discret si dificil de pus in evidenta
la alte specii - gros, aceste substante mucoase sunt secretate in cantitati excesive
Capsula
un invelis extraparietal bine organizat si individualizat morfo-chimic, care se formeaza numai cand bacteriile ajung in conditii favorabile sintezei acestuia, fiind o caracteristica determinata genetic
B. anthracis si Str. pneumoniae
- numai cand germenii au patruns in organism si vin in contact cu umorile acestuia;
- in natura sau in culturile dezvoltate pe mediile uzuale, germenii sunt obisnuit necapsulati.
Sinteza este supusa variabilitatii:
variante capsulogene - capabile sa sintetizeze capsula in orice conditii de mediu (B. anthracis)
variante acapsulogene care au pierdut definitiv (genetic) proprietatea de a capsula
Capsula formeaza in jurul celulei un invelis continuu, cu aspect aparent omogen
Afinitati tinctoriale distincte de cele ale peretelui bacterian
Componentele care intra in structura sa sunt diferite chimic si antigenic, de la o specie la alta:
numai componente glucidice (polizaharide capsulare)
numai componente proteice;
complexe proteice si glucidice.
Rol biologic
protector - bacteriile saprofite - capsula stanjeneste inglobarea lor de catre protozoare.
in virulenta - la bacteriile patogene capsula indeplineste functii deosebit de complexe in procesul de agresiune, fiind considerata unul din factorii de patogenitate
antifagocitar
imposibila fagocitoza, intrucat bacteriile capsulate nu mai adera la membrana fagocitelor si, ca urmare, nu pot fi inglobate de aceste celule
un efect de paralizare a fluxului leucocitar spre focarul de infectie, germenii patogeni se pot multiplica in umorile si tesuturile in care au patruns, realizand pana la urma invazia organismului.
ORGANITE CELULARE
Cili (Flageli)
Pili (fimbrii)
Cilii (Flagelii)
sunt organite cilindrice foarte subtiri, lungi si flexibile, cu numeroase ondulatii helicoidale regulate
cu ajutorul carora pot efectua in mediile hidrice diferite deplasari taxice, cu viteze relativ mari
existenta cililor, numarul si pozitia lor pe suprafata bacteriei - determinate genetic
de mare importanta in taxonomie si in lucrarile de diagnostic
filamentul ciliar este format predominant din molecule proteice sferoidale denumite flagelina (antigenica - antigenii "H" ai bacteriilor mobile)
ei pot fi pusi in evidenta prin metode speciale de colorare
in lucrarile de rutina se recurge numai la evidentierea indirecta, prin examenul mobilitatii bacteriilor
Tipuri:
atricha (gr. tricha = par) - lipsite de cili si deci imobile;
monotricha - un singur cil sau flagel situat la un pol al celulei;
amphitricha - doi cili situati cate unul la polii celulei;
lophotricha - mai multi cili (5-30) grupati intr-un smoc situat la un pol al celulei;
peritricha - un numar mare de cili (intre 30-100 cili) dispusi jur-imprejur, pe toata suprafata celulei (Proteus vulgaris numarul lor ajunge chiar la cateva sute)
Infrastructura cililor este relativ complexa, distingandu-se trei componente:
corpusculul bazal
carligul sau articulatia
filamentul ciliar helicoidal
Corpusculul bazal
este formatiunea cu care cilii se ancoreaza in membrana citoplasmatica si in stratul profund al peretelui bacterian (stratul "R" sau "rigid")
indeplineste rolul de piesa "rotor", capabila sa imprime cililor miscari rotatorii intr-un sens sau altul
in structura sa se disting 4 discuri la bacteriile Gram-negative si numai doua discuri la bacteriile Gram-pozitive
Ele sunt denumite prin initialele dupa rolul pe care il au sau stratul parietal in care se gasesc:
discul "M" (de membrana) - in stratul dublu lipidic al membranei citoplasmatice - rol de rotor;
discul "S" sau stator - fiind situat la fata externa a membranei citoplasmatice;
discul "P" este inclavat - in stratul mureinic rigid ("R");
discul "L" - in profunzimea membranei externe lps a peretelui.
Discurile sunt interconectate printr-o piesa intermediara care indeplineste rolul de ax de transmisie a miscarilor rotatorii.
Carligul (sau articulatia) este o componenta flexibila, intercalata intre corpusculul bazal si filamentul ciliar
Filamentul ciliar helicoidal reprezinta cilul propriu zis - din molecule sferoidale de flagelina dispuse in aranjament helicoidal.
Rol biologic
permite bacteriilor sa efectueze in mediile hidrice numeroase deplasari taxice, orientate spre zonele care le ofera conditii optime de viata (bacteriile aerobe se deplaseaza spre zonele in care oxigenul se gaseste in concentratie maxima - aerotaxie)
miscarile rotatorii si ondulatorii sunt executate sincronizat, in acelasi sens de toti cilii, ceea ce imprima celulei o deplasare orientata in spatiu intr-o anumita directie.
unele specii isi pot sincroniza miscarile ciliare la toti germenii dintr-o colonie, ceea ce poate imprima deplasarea lenta a acesteia pe suprafata mediilor semilichide.
Pilii (fimbriile)
Sunt organite filamentoase, cilindrice, foarte subtiri, scurte si drepte, aparent rigide, dispersate pe toata suprafata celulei (ca o perie)
Pilii pot fi pusi in evidenta numai la unele bacterii Gram-negative, prin microscopie electronica.
Pe baza unor caractere morfo-functionale se diferentiaza in 6 tipuri: I, II, III, IV, V si F (pilii de sex).
Structura morfo-chimica
Pilii de tip I-V sunt organite tubulare subtiri, formate din molecule de fimbrilina, dispuse pe un singur rand, intr-un aranjament helicoidal .
Pilii de tip F sau pilii sexuali sunt organite tubulare putin mai groase si mai lungi, formati din molecule de pilina, o fosfoglicoproteina cu masa moleculara de 11.800 daltoni.
subunitatile acesteia se dispun, pe un singur rand, intr-un aranjament helicoidal, delimitand un canal axial, prin care pot trece moleculele de ADN dublu catenar
Rol biologic
Pilii de tip I-V fac posibila fixarea bacteriilor pe anumite suporturi celulare sau de alta natura pe care le pot astfel si coloniza.
Pilii F (sexuali) sunt organite care asigura in cursul proceselor de conjugare, cuplarea celulelor bacteriene F+ si F-, diferentiate ca polaritate sexuala, canalul piliar servind la transferul de material genetic (ADN) de la celula F+ la F-.
Pilii F (de sex) se gasesc la suprafata celulelor F+. Sinteza lor este specificata si controlata de plasmida F (sau de alte plasmide "sexductoare").
SPORUL BACTERIAN
este o formatiune morfo-biologica dotata cu rezistenta crescuta la agentii fizico-chimici prin care bacteria isi poate conserva viabilitatea timp indelungat, in stare latenta, in conditii in care forma vegetativa nu ar putea supravietui
Proprietatea de a sporula o au un numar restrans de specii bacteriene, toate incadrate in doua familii:
fam. Bacillaceae care incadreaza speciile aerobe ce sporuleaza facultativ;
fam Clostridiaceae care incadreaza speciile anaerobe ce sporuleaza constant.
Sporogeneza
Sporul se formeaza in celula bacteriana prin procese complexe de reorganizare morfo-chimica.
De regula, dintr-o celula bacteriana rezulta un singur spor, iar din acesta - prin germinare - ia nastere o singura forma vegetativa
Ca urmare, sporogeneza nu constituie pentru bacterii o modalitate si un mijloc de multiplicare.
Sporogeneza este considerata ca un fenomen normal in evolutia celulei bacteriene, procesul fiind controlat si dirijat morfogenetic de cca 50 de gene
celula devine competenta sa sporuleze numai cand atinge in evolutia sa un anumit stadiu, o anumita maturitate biologica.
formele vegetative noi apar intinerite, fiind foarte active metabolic si avand un ritm de diviziune deosebit de intens
Sporularea si germinarea sporilor nu sunt fenomene intamplatoare, ci procese supuse permanent unor mecanisme de autocontrol si reglare genetica, dar care pot fi induse de numerosi factori endogeni si exogeni.
Sporogeneza si germinarea sporilor sunt procese controlate si dirijate genetic de aproximativ 50 de gene, situate in cromozom in locus-uri diferite.
Acestea se mentin in forma vegetativa in stare represata, pana cand sub actiunea unor factori inductori endogeni si exogeni (de mediu) sinteza represorului inceteaza.
Factorii de mediu exercita un rol activant sau inhibant asupra sporogenezei, cat si asupra germinarii sporilor,
procesul de germinare este influentat pozitiv de o temperatura apropiata de cea optima multiplicarii, de umiditate, de diferite substante prezente in mediu (glucoza, acizi aminati ca L-alanina, ioni de Na, K si Ca etc.).
De exemplu, B. anthracis sporuleaza la temperaturi cuprinse intre 16°-42°C; in afara acestor limite sporularea inceteaza, desi multiplicarea germenilor mai poate continua.
Oxigenul molecular este un factor indispensabil sporularii bacteriilor aerobe; pentru bacteriile anaerobe este insa nociv, sporularea lor putand fi inhibata complet de prezenta acestuia in mediu.
Rapiditatea de sporulare variaza, de altfel, chiar de la o celula la alta. Pe de alta parte, proprietatea de a sporula poate fi pierduta, putandu-se obtine variante asporogene stabile genetic.
Structura morfo-chimica
Sporii au o forma sferoidala, usor ovalara, dimensiunile variind cu specia intre 0,5-2 µm.
Se pot forma in celula bacteriana in:
zona centrala,
subterminala (spre un capat)
terminala (la capatul formei vegetative)
Dimensiunile si pozitia sporilor, raportate la forma vegetativa, sunt caracteristice la unele specii bacteriene si au valoare taxonomica, fiind utile in lucrarile de identificare a genului si a speciilor:
sporii speciilor din genul Bacillus au
diametrul mai mic decat grosimea bacililor,
se formeaza in zona centrala sau subterminala
nu imprima deformarea formei vegetative
La speciile din genul Clostridium, diametrul sporilor depaseste, de regula, grosimea formei vegetative, imprimand deformarea acesteia;
cand sporii ocupa o pozitie centrala, bacteriile sporulate iau o forma naviculara, asemanatoare cu o suveica (bombate in zona centrala),
cand sporii au o pozitie subterminala, bacilii iau forma bombata spre unul din capete, asemanatoare cu a unei sticle de lampa sau cu o para.
La Clostridium tetani sporii sunt, de regula sferoidali, se formeaza in pozitie terminala si raman atasati la forma vegetativa, dand bacililor un aspect caracteristic de "ciocanas de batut toba" sau de "racheta de tenis".
Aspecte similare sunt intalnite la Cl. tetanomorfus, dar sporii sunt mult mai mari si au o forma pronuntata ovalara
Infrastructura este asemanatoare la toti sporii.
Protoplastul sporal (celula sporala propriu-zisa) contine:
materia nucleara (sau nucleoplasma) formata din ADN cromozomal
o cantitate relativ redusa de sporoplasma cu aspect granular (probabil ribosomii), mult mai densa si electroopaca decat citoplasma.
Acest continut sporal este delimitat de o membrana sporala, care constituie totodata stratul intern al peretelui sporal pluristratificat.
Ansamblul acestor caracteristici structurale fac ca sporii sa aiba afinitati tinctoriale diferite de ale formelor vegetative.
Ei pot fi colorati numai prin metode energetice, care forteaza patrunderea colorantilor prin perete si in continutul sporal.
Sporii se deosebesc fundamental de formele vegetative prin numeroase alte caracteristici fizico-chimice.
Contin foarte putina apa libera, putine enzime si o cantitate mult mai mica de ribonucleoproteine
sunt mai saraci in saruri de K si P, dar mai bogati in ioni de Ca, Mg si Mn, si de 4-5 ori mai bogati in cisteina.
contin o cantitate apreciabila de acid dipicolinic (existent numai in spor, in cantitate de cca 10% din greutatea uscata). Acesta se gaseste sub forma de sare de Ca, provine probabil din ac. diaminopimelic si are - se pare - rol in conferirea termostabilitatii enzimelor sporale, la care se leaga si le da o configuratie aparte.
Sporii nu desfasoara o activitate metabolica si nu au schimburi de substante cu mediul.
Enzimele pe care le contin in cantitate redusa sunt sub o forma inactiva.
In schimb, contin enzime catalitice fata de alanina in cantitate de 2-10 ori mai mari decat formele vegetative, acestea avand un rol important in procesele de germinare.
Rezistenta la factorii fizico-chimici
Sporii au rezistenta mare la numerosi agenti fizico-chimici.
Se remarca in special rezistenta termica mare
sporii fiind distrusi la caldura umeda Ia temperatura de 120°C, dupa 20-30 minute, iar la caldura uscata de 160-180°C dupa 30-60 minute
sporii unor specii bacteriene pot rezista la fierbere pana la 1-2 ore, in timp ce formele vegetative mor obisnuit in 10 minute la 70-80°C.
Rezistenta sporilor este deosebit de mare la uscaciune si la radiatii.
Ei se conserva si isi pot mentine viabilitatea in pamant si in diverse materiale patologice, timp foarte indelungat.
sporii de B. anthracis pot ramane viabili in pamant timp de peste 70 de ani,
sporii unor bacterii saprofite si-au pastrat viabilitatea in conserve alimentare timp de peste 114 ani
la fel se comporta si sporii de Cl. tetani
In comparatie cu formele vegetative, sporii sunt mult mai rezistenti la substantele antiseptice si la dezinfectante.
sporii bacililor carbunosi se mentin viabili cativa ani in solutie cu adaos de 40-50% glicerina, in timp ce formele vegetative sunt distruse in cateva ore
sporii unor bacterii raman vii pana la o saptamana in solutie de fenol 5% si mai multe saptamani in alcool.
Germinarea sporilor
Transformarea sporilor in forme vegetative este un proces complex, care se desfasoara in stransa corelatie cu factorii de mediu, in trei etape succesive
activarea,
germinarea
cresterea formei vegetative
noi
Activarea si germinarea sunt fenomene stimulate de caracterul hidric al mediului si de prezenta unor componente chimice usor metabolizabile.
Procesul consta intr-o reorganizare rapida a componentelor sporale.
In prima faza se produce o absorbtie intensa si rapida de apa, insotita de cresterea in volum a sporului uneori de 100% si de pierderea refringentei.
Concomitent, se petrec modificari structurale si biochimice, in cursul carora se produce activarea si imbogatirea echipamentului enzimatic cu rol energetic si plastic.
Multe componente sunt degradate si restructurate in componente noi, unele substante specifice sporului fiind eliminate (acidul dipicolinic, anumite peptide, saruri de Ca etc.).
Nucleul revine in pozitie centrala, iar in cortexul sporal se produc schimbari esentiale, finalizate la scurt timp prin ruperea peretelui sporal si prin aparitia unei emergente care creste rapid, dand nastere unei noi forme vegetative. Aceasta se desprinde, de regula, de membrana sporala, dar uneori invelisul sporal ramane atasat acesteia o anumita perioada.
Viteza de germinare difera in limite largi, in functie de specie si de vechimea sporilor.
De exemplu, sporii de B. anthracis recent formati, ajunsi in conditii favorabile, germineaza de regula in 60-90 secunde, in timp ce germinarea sporilor vechi incepe abia dupa 1-2 ore sau chiar mai mult.
FIZIOLOGIA BACTERIANA
Pe parcursul evolutiei lor s-au adaptat la conditii de mediu si viata mult mai variate decat lumea vegetala.
Ca urmare, si sub aspect metabolic lumea bacteriana este mai putin omogena, fiind deosebit de diversificata mai ales in privinta capacitatii de a utiliza componentele existente in mediu ca surse nutritive.
S-au conturat astfel numeroase grupe specializate de bacterii, cu caracteristici ecologice si metabolice foarte variate, care le permit sa indeplineasca in natura un rol esential in circulatia complexa a materiei, in sensuri multiple.
Cunoasterea caracteristicilor metabolice are o deosebita importanta practica:
in cultivarea bacteriilor pe medii artificiale,
in lucrarile de diagnostic,
in selectarea de tipuri cu proprietati fermentative utile in diferite ramuri industriale etc.
STRUCTURA CHIMICA A BACTERIILOR
Celula bacteriana, are o structura chimica deosebit de complexa.
in structura celulei bacteriene se gasesc toate componentele caracteristice materiei vii, bacteriile isi au totusi individualitatea lor chimica, deosebindu-se de alte vietuitoare mai ales prin unele componente chimice macromoleculare care sunt proprii lor.
Aproape jumatate sunt
componente anorganice cu molecula mica (H2O, saruri minerale);
componente organice moleculare si macromoleculare;
- glucide complexe si precursori (cca 200 tipuri),
- lipide (cca 50 tipuri),
- proteine (106 molecule care apartin la cca 2000-3000 tipuri) etc.
Structura chimica elementara
In structura celulei bacteriene intra toate cele 11 elemente biogene caracteristice materiei vii: C, H, O, N, P, Ca, Na, Cl, S, Mg, K.
Cantitativ acestea reprezinta 99,9% din totalul elementelor. Restul de 0,1% sunt oligoelemente: Cu, Co, I, Br, Bor etc.
Cele mai multe intra in structura unor enzime si factori de crestere.
In comparatie cu alte vietuitoare, bacteriile sunt in general mai bogate in C si N (in concordanta cu bogatia lor in proteine), dar sunt mai sarace in O si Ca.
Contin cantitati ceva mai mari de P, Na si Mg.
Continutul in elemente chimice variaza insa, in anumite limite, de la o specie la alta, iar la aceeasi specie in functie de mediul de cultivare, de starea fiziologica si de varsta celulelor bacteriene etc.
Multe componente se leaga intre ele formand complexe macromoleculare caracteristice, fie cu rol structural (mureina, lps etc.), fie cu rol functional (enzime, exotoxine etc.).
Apa
Este componentul predominant cantitativ.
Reprezinta aproximativ 75-85% din greutatea umeda a celulei bacteriene.
O parte se gaseste sub forma de apa de constitutie, incoporata in structura diverselor componente chimice celulare.
Cea mai mare parte se gaseste insa ca apa libera, conferind mediului endocelular un pronuntat caracter hidric.
In apa libera se gasesc solvate
- cantitati apreciabile de saruri minerale
- componente metabolizabile sau rezultate din activitatea metabolica.
De caracterul hidric endocelular depinde desfasurarea in bune conditii a intregului metabolism
- realizarea schimburilor active si continui dintre celula si mediu,
- difuziunea enzimelor in mediul endocelular si spre exterior,
- eliminarea metabolitilor,
- patrunderea si difuziunea componentelor utile in metabolismul plastic si energetic etc.)
Totodata, apa libera participa ca substrat in majoritatea reactiilor biochimice (hidroliza, oxidoreducere etc.).
Diminuarea continutului in apa determina o incetinire a ritmului de desfasurare a proceselor metabolice.
Apa intervine in reglarea echilibrului osmotic endocelular, a pH-ului si a potentialului oxidoreducator.
Continutul in apa libera influenteaza apoi rezistenta celulei la unii factori chimici si fizici, diminuarea ei crescand rezistenta bacteriei la uscaciune, temperatura si la unele substante chimice.
Sarurile minerale
Continutul in saruri minerale este cuprins intre 2-30% din greutatea uscata a celulei bacteriene.
Cantitatea acestora variaza in raport cu specia, cu starea fiziologica a celulei, cu compozitia chimica a mediului etc.
De regula, celulele tinere contin saruri minerale in cantitati de 6-7 ori mai mari decat celulele batrane.
Dintre componentele minerale cele mai bine reprezentate cantitativ sunt: P, K, Na, Mg, Ca, S, Cl. In cantitati mai mici se gasesc Zn, Fe, Cu, Mn etc.
Predomina compusii de P, care reprezinta in medie 45% din totalul componentelor minerale (la M. tuberculosis pot ajunge pana la 74%).
Unii compusi minerali intra in structura componentelor macromoleculare cu rol plastic;
altii intra in structura enzimelor si coenzimelor sau indeplinesc rol de activatori in diverse procese metabolice (Ca, Na, Zn).
Constituentii minerali intervin apoi in numeroase alte procese: asigura reglarea presiunii osmotice endocelulare si mentinerea echilibrului de membrana, influentand permeabilitatea acesteia; contribuie la reglarea pH-ului si a potentialului oxido-reducator etc.
Compusii de P au rol esential in metabolismul energetic al celulei bacteriene. In legaturile fosforice macroergice ale acestora se pot stoca cantitati mari de energie, pe care celula o poate elibera in raport cu cerintele sale.
Bacteriile sunt vietuitoare care au necesitati minerale mari si multiple.
Cunoasterea acestor cerinte are o deosebita importanta in practica cultivarii germenilor pe medii artificiale, iar in multe cazuri chiar la izolarea si identificarea unor specii.
Glucidele
cantitate mare si variata de glucide (ozide si poliozide), acestea reprezentand intre 4-25% din greutatea uscata a celulei.
Continutul lor variaza insa cu specia si starea fiziologica
Glucidele au rol biologic complex, o mare cantitate fiind utilizata de celula bacteriana ca surse de carbon si energie.
Numeroase glucide formeaza complexe macromoleculare caracteristice care intra in structura celulei bacteriene, in special in structura peretelui bacterian si a capsulei
O parte din glucide pot fi polimerizate si depozitate in citoplasma unor bacterii ca materii de rezerva, sub forma de granule de amidon si glicogen.
Ozidele se gasesc fie libere in curs de metabolizare, fie legate de componente de alta natura.
Cele mai frecvente sunt: glucoza (sau derivati ca: glucozamina, acidul glucuronic), galactoza, manoza, arabinoza etc.
Dintre pentoze, riboza si dezoxiriboza intra in structura acizilor nucleici.
In celula bacteriana pot fi gasiti frecvent polimeri complecsi formati din ozide conjugate cu acizi uronici: acidul hialuronic care intra in structura capsulei la streptococi, polizaharidele capsulare la pneumococi etc.
Unele glucide se leaga cu anumite polipeptide formand polimeri complecsi cu rol structural, cum este, de exemplu, mureina din structura peretelui bacterian.
Alte glucide formeaza cu diverse lipide complexele lipopolizaharidice (lps) prezente in peretele celular la bacteriile Gram-negative, constituind antigenii "O" si endotoxinele acestora.
Multe polizaharide extrase din celula bacteriana au caracter de antigeni incompleti sau haptene (pot intra in reactie cu anticorpii, dar nu pot induce sinteza lor).
In celula acestea se gasesc insa legate de alte componente, de regula proteice, cu care formeaza antigeni complecsi, glucidele avand rolul de grupari active purtatoare ale specificitatii antigenice).
Lipidele
Difera cantitativ in limite largi de la o specie la alta, ele reprezentand intre 2-40% din greutatea uscata a celulei.
Bacteriile cele mai sarace in lipide par sa fie stafilococii (2,8%), iar cele mai bogate in lipide bacteriile din genul Mycobacterium.
Continutul in lipide variaza la aceeasi specie in functie de varsta celulelor si de compozitia chimica a mediului.
La unele specii bacteriene lipidele se pot acumula in celula si ca materie de rezerva, mai ales cand germenii sunt cultivati in conditii restrictive (pe medii bogate in C si in surse energetice, dar sarace in N) sau pe medii cu o anumita compozitie chimica.
Lipidele bacteriene sunt in general foarte variate, o parte gasindu-se sub forma de compusi esterificati cu alcooli.
Multe lipide reprezinta pentru celula bacteriana o importanta sursa de C si energie.
Unele indeplinesc rol structural, cum sunt cele din peretele bacteriilor Gram-negative si din peretele bacteriilor acido-rezistente
Fosfo-lipidele intra in structura membranei citoplasmatice.
In general, lipidele bacteriene pot fi grupate in trei categorii:
a) lipide neutre, unele acumulandu-se in celula ca materie de rezerva, sub forma de polimeri ai acidului betahidroxibutiric;
b) ceride, formate prin esterificarea acizilor grasi cu alcooli. Sunt continute mai ales de bacteriile acido-rezistente in proportii de 5-10% din greutatea lor uscata. Lipsesc sau se gasesc in cantitati reduse la bacteriile neacido-rezistente;
c) fosfolipidele, bine reprezentate cantitativ la toate bacteriile (contin acizi grasi, alcooli, baze azotate si acid fosforic).
De exemplu, la enterobacterii fosfolipidele reprezinta pana la 50% din totalul lipidelor; lipidele din complexele lps sunt, de asemenea, fosfolipide in proportie de 80-95%.
Proteinele
Bacteriile sunt foarte bogate in proteine, acestea reprezentand in medie 60% din greutatea lor uscata.
Se apreciaza ca intr-o celula exista cca 106 molecule proteice, apartinand la 2000-3000 tipuri.
Variatiile cantitative si calitative sunt in general restranse, dependente mai ales de starea fiziologica a celulelor bacteriene.
O mare parte a componentelor proteice intra
- in structurile celulare,
- in stare de dispersie coloidala in mediul citoplasmatic,
- fie sub forma unor componente in
curs de metabolizare,
- fie ca proteine cu rol functional
(enzime, exotoxine).
se poate afirma ca personalitatea chimica si imunologica a bacteriilor este data in special de componentele proteice.
sinteza proteinelor este controlata genetic si decurge conform informatiei inscrise in ADN-ul fiecarei specii bacteriene,
|
Politica de confidentialitate |
| Copyright ©
2025 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Apa in organism |
| MORFOLOGIE BACTERIANA |
| CAILE RESPIRATORII |
| CAVITATEA TORACICA SI CAVITATILE PLEURALE |
| MEMBRANA CELULARA |
| TRANSMITEREA INFORMATIEI EREDITARE |
| Increngatura Mollusca |
| Arterele |
| Termeni si conditii |
| Contact |
| Creeaza si tu |
