
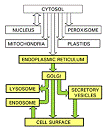
Organitele celulare cu rol in transportul și selectarea intracelulara a moleculelor
Reticulul endoplasmatic
Aparatul Golgi
Mecanismul transportului vezicular
Lizozomii
Reticulul endoplasmic (RE) este un organit delimitat de endomembrana, structurat sub forma unor cisterne și/sau tubuli, cu numeroase anastomoze, a caror fața citoplasmatica prezinta, sau nu, rugozitați și a carui funcție de baza este aceea de a produce molecule și macromolecule esențiale organizarii și funcționarii celulelor.
RE face parte din grupul organitelor implicate in biogeneza și traficul intracelular al membranelor, alaturi de aparatul Golgi, lizosomi și sistemul endosomal, fiind primul din serie, adica cel care inițiaza procesele celulare care se desfașoara in aceste organite.
Reticulul endoplasmic reprezinta cea mai abundenta structura delimitata de endomembrane din celula, conținand mai mult de jumatate din membranele acesteia.
Exista doua tipuri de RE: RE rugos (RER) care este acoperit de ribozomi pe suprafața exterioara și are rolul de prelucrare a proteinelor.
REN (neted) care nu este asociat cu ribozomii fiind implicat mai ales in metabolismul lipidic.
RE si sinteza proteinelor
Funcțiile RER(rugos)
1. Biosinteza unor proteine: (i) proteine membranare, (ii) proteine destinate a funcționa in RE, in aparatul Golgi, sau lizosomi, (iii) proteine destinate exportului;
2. Prelucrarea proteinelor sintetizate in RE;
3. Sortarea și transportul catre aparatul Golgi.
Biosinteza proteica la nivelul RE
Biosinteza tuturor proteinelor intr-o celula se inițiaza in citosol. Excepție fac proteinele codificate de ADN-ul mitocondrial (puține; nu mai mult de 10% dintre proteinele necesare funcționarii mitocondriei).
Se pune problema: cum știe RE care dintre complexele de biosinteza proteica (polisomi) trebuie sa fie preluate la nivelul membranei sale? Informația prin care se face selecția se afla in insuși lanțul polipeptidic in formare. Ea este o secvența compacta de 15-30 aminoacizi hidrofobi .
In 1960,George Emil Palade (premiul Nobel 1974 pt. descoperiri in domeniul biomedical) și colaboratori au demonstrat pentru prima oara rolul RE in sortarea și prelucrarea proteinelor. Calea secretorie a proteinelor este: RER- Ap. Golgi- Vezicule secretorii- exteriorul celulei.
Exista doua tipuri de transport prin care circula proteinele sintetizate la nivelul ribozomilor:
Transportul post-translațional - proteinele destinate sa ramana in citosol sau sa fie incorporate in nucleu, mitocondrii, peroxizomi sunt sintetizate in ribozomii liberi și eliberate in citosol cand translația lor s-a terminat, deci sinteza e finalizata.
Transportul co-translational - majoritatea proteinelor destinate secreției sau incorporarii in RE, ap. Golgi, lizozomi sau membrane plasmatice, sunt sintetizate in ribozomii legati la membrana și transportate in RER pe masura ce translația are loc.
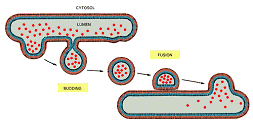
Mecanismele fundamentale de deplasare a proteinelor de la un compartinent la altul
Transport prin translocatori proteici= mecanism de transport care se face in mod direct, prin bistratul lipidic, cu ajutorul unor proteine inserate in membranele care separa cele 2 compartimente.
Transportul prin vezicule- veziculele inglobeaza macromolecule din lumenul unui compartiment desprinzandu-se prin "inmugurire" de membrane acestuia, pentru ca apoi sa-și descarce conținutul in lumenul altui compartiment, ca urmare a fuziunii cu acesta. Nu strabat nici o membrana.
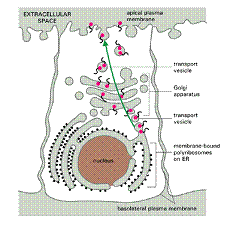
Orientarea proteinelor de secreție catre RE este realizata de secvențele semnal care au 15-30 aminoacizi, reprezinta o porțiune intinsa de resturi hidrofobe. Pe masura ce ies din ribozomi, secventele semnal sunt recunoscute și legate de o particula de recunoaștere a semnalului (SRP). Particulele SRP sunt macromolecule complexe, formate din proteine și ARN.
Receptorul pentru SRP care este inserat in membrana RE si leaga specific particulele de SRP , in momentul in care ele s-au atașat la peptidul semnal de pe lanțul polipeptidic in crestere.
Legarea de receptor elibereaza SRP atat de ribozom cat și de secvența semnal a lanțului polipeptidic care se naște.
Ribozomul, in continuare se leaga la un complex de translocare proteic (dim membrana RE) , secventa semnal este introdusa intr-un canal membranar. Translația se poate relua, iar lanțul polipeptidic in creștere este translocat prin membrana in RE.
Pe masura ce translocarea are loc, secvența semnal este clivata de o peptidaza semnal și polipeptidul este eliberat in lumenul RE.
Proteinele care vor fi incorporate in membrana plasmatica sau in membranele RE, ap. Golgi sau lizozomilor sunt inițial inserate in membrana RE fara sa fie eliberate in lumen. De la membrana RE ele circula, catre destinația finala ca și proteine de secreție pe aceeasi cale (RE-Ap. Golgi- membrana plasmatica sau lizozomi) . Transportul lor se face sub forma de componente membranare si nu proteine solubile.
REN și sinteza lipidelor
Este portiunea cea mai importanta pentru sinteza lipidelor.
Deoarece sunt extrem de hidrofobe, lipidele nu sunt sintetizate in mediul apos al citosolului, ci in asociere cu alte membrane in RE, apoi sunt transportate din RE catre destinațiile finale, fie in vezicule, fie prin proteine carrier.
La acest nivel sunt sintetizate fie formele finale fie precursori ai lipidelor. Cele mai multe lipide sunt de tipul fosfolipide, glicolipide, colesterol. Cele mai multe deriva din glicerol si sunt sintetizate pe fata citosolica a membranei RE din precursori citosolici.
Noile fosfolipide vor fi sintetizate numai pe fața membranara citosolica, de unde sunt translocate cu ajutorul unei proteine denumite flipaze, catre jumatatea luminala a bistratului, ceea ce echilibreaza distribuția fosfolipidelor.
Colesterolul este produs in RE printr-un proces biologic complex, bine elaborat și atent
reglat, format din multe etape. Materia prima este acetil-CoA (CoA - coenzima A), iar intermediarul de baza este acidul mevalonic, format prin activitatea HMG CoA reductazei (HMG - 3-hidroxi-3metilglutaril).
In etapele urmatoare, prin intermediul farnezil-fosfatului se produce scualenul, sub acțiunea scualen-sintazei, care apoi sufera, sub acțiunea scualen-oxidociclazei, ciclizarile ce duc la obținerea intermediarului conținand nucleul tetraciclic, lanosterolul. Transformarea lanosterolului la cholesterol implica multe faze mai puțin elucidate. Enzimele menționate mai sus fac toate parte din bagajul molecular al RE.
Tot la nivelul RE sunt produse ceramidele, precursorii sfingomielinelor și glicolipidelor. Ceramidele se obțin prin amidarea sfinganinei, un aminodiol alifatic, precursor al sfingozinei obținut din L-serina și palmitil-CoA. Dihidro-ceramidele astfel obținute sunt dehidrogenate. Ceramidele sunt transformate in sfingomieline, sau glicolipide (cerebrozide) la nivelul complexului Golgi.
Un aspect interesant, care merita punctat este faptul ca, celula nu este nevoita sa sintetizeze fosfolipidele de novo, atunci cand proporția dintre diferitele tipuri trebuie sa se schimbe la nivelul bistratului. Fosfolipidele pot suferi reacții de disproporționare, adica acele reacții prin care ele pot trece dintr-una in alta. Posibilitațile de disproporționare nu sunt nici universale (adica oricare dintre ele sa poata trece in oricare dintre celelalte), nici intotdeauna bidirecționale. Astfel sunt cunoscute urmatoarele posibilitați de disproporționare:
a) La nivelul RE:
1. fosfatidiletanolamina poate trece in fosfatidilcolina (conversia implica reacții de metilare pentru care exista enzima adecvata:
fosfatidiletanolamin-N-metil-transferaza
2. exista posibilitați de conversie in ambele sensuri intre fosfatidilcolina, respectiv fosfatidiletanolamina și fosfatidilserina (prin reacții de schimb la nivelul capului hidrofil: colina, sau etanolamina sunt schimbate cu serina, sub acțiunea unor fosfatidilserin- sintaze); de menționat ca PS se produce numai prin acest mecanism de schimb in celulele de mamifere.
b) La nivelul mitocondriei
1. fosfatidilserina poate trece in fosfatidiletanolamina (prin decarboxilare sub
acțiunea fosfatidilserin-decarboxilazei)
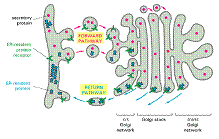
Exportul proteinelor și lipidelor din RE
Moleculele sunt exportate in vezicule formate prin inmugurire din RE. Apoi veziculele fuzioneaza cu membrana aparatului Golgi și de aici pot sa ajunga la lizozomi sau la membrana plasmatica.
In timp ce sunt transportate prin vezicule, lipidele si proteinele, nu-si modifica orientarea spatiala. Daca trebuie sa ramana in RE proteinele sunt marcate prin secvente tinta care impiedica impachetarea si transportarea lor in vezicule.
Aspecte legate de metabolismul lipidelor
Un alt proces care implica metabolismul lipidic, cu importanța in capacitatea celulelor de a modula proprietațile fizico-chimice ale membranelor, este desaturarea acizilor grași Aceasta se face prin acțiunea unui complex enzimatic ce conține citocrom B5, NADH-citocrom B5-reductaza și acid gras desaturaze.
Procesul are loc adesea cu alungirea lanțului alifatic. Nu exista dovezi ca aceste procese s-ar petrece direct pe fosfolipide, ci doar pe acizii grași esterificați, ca tioesteri, cu CoA. Modularea cantitații de acizi grași nesaturați in fosfolipidele membranare permite celulelor sa-și regleze fluiditatea membranelor, in conformitate cu nevoile de moment. Faptul ca desaturarea se face pe acizi grași in afara lipidelor membranare ar insemna ca modularea fluiditații se face prin sinteza de novo a fosfolipidelor. O problema care se ridica este legata de eficiența raspunsurilor in modularea fluiditații pe aceasta cale.
Detoxificarea celulara
Procesele care rezolva aceasta problema implica metabolizarea, pentru eliminarea din celula, a compușilor liposolubili, care s-ar putea acumula in bistratul lipidic, afectandu-i fluiditatea intr-un mod necontrolat de celula.
Acești produși pot fi fiziologici, patologici, sau farmacologici. La nivelul RE acești produși hidrofobi sunt mai intai hidroxilați prin acțiunea unui complex enzimatic bazat pe citocrom P450/NADPH-citocrom P450-reductaza. Ei sunt astfel transformați in structuri hidrofile, ușor de eliminat din celula. Daca este cazul, aceste prime modificari sunt urmate de grefarea, la gruparile hidroxil astfel obținute, a unor structuri glucidice sau grupari sulfat, care maresc hidrofilicitatea produșilor rezultați.
Rolul in detoxificarea celulara este spectaculos sugerat și de fenomenul de hiperplazie (creșterea cantitații, sau numarului de structuri) a REN in hepatocitele indivizilor medicați, pentru o perioada mai indelungata, cu barbiturice.
La scurt timp dupa inceperea perioadei de tratament, crește semnificativ cantitatea de REN in celulele ficatului. Hiperplazia este reversibila, cantitatea de REN revenind la normal la scurt timp dupa incetarea medicației. De remarcat faptul ca, in hepatocitele normale, structurile de RE prezinta o proporție de echilibru (~1:1) intre RER și REN, astfel incat modificarile acestui raport sunt ușor de observat.
Reticulul endoplasmic - depozit dinamic de Ca2+
Aceasta funcție este pregnant manifesta la celulele musculare striate. La aceste celule, la care reticulul endoplasmic este denumit reticul sarcoplasmic (RS), funcția și dinamica ei sunt realizate prin cooperarea mai multor componente moleculare. O prima componenta este calsechestrina, proteina cu mare afinitate pentru ionii de calciu, aflata in cantitate mare in lumenul organitului.
Prezența calsechestrinei contribuie la controlul cantitații de Ca2+ liber din lumen (conform constantei sale de afinitate), in condițiile unei concentrații totale de Ca2+ crescute. La stimularea celulelor, se deschid in membrana RS canale de calciu controlate chimic (prin inozitol tris-fosfat - IP3), prin care ionii de calciu, aflați liberi in lumen, patrund in citosol și declanșaza contracția.
Trecerea Ca2+ din lumenul RS in citosol are
loc atata timp cat canalele sunt deschise, pe baza deplasarii echilibrului
dinspre calciul legat pe calsechestrina, spre calciul liber. Ciclul se inchide
prin acțiunea unor pompe de calciu din membrana RS, care reintroduc
Ca2+ in lumenul RS, unde calsechestrina il complexeaza, pentru a
pastra
(vezi curs 5- transport membranar)
Abundența și distribuția intracelulara a RE
Reticulul endoplasmic este un organit ubicuitar. Rolul sau in biogeneza membranelor il face indispensabil organizarii și funcționarii celulelor. Chiar și in cazul eritrocitului (lipsit de organite), reticulul endoplasmic a fost prezent și a activat in timpul diferențierii precursorilor, pana in momentul maturarii elementului circulant. Daca, deregula, RE conține cel puțin jumatate din membranele dintr-o celula, raportul dintre componenta rugoasa și cea neteda variaza in funcție de tipul de celula.
Exista celule in care RER este preponderent (celule specializate in sinteza și secreția de proteine; exemplul tipic il formeaza celulele acinare pancreatice), sau celule in care REN este
preponderant (celule specializate in sinteza și secreția de hormoni steroidici; de exemplu celulele zonei corticale a glandei suprarenale, sau celulele Leydig din testicul).
Un alt caz (reprezentat prin hepatocite de exemplu) este acela al celulelor in care raportul
RER/REN este echilibrat. Cat privește distribuția intracelulara a RE aceasta poate fi difuza, cum ar fi in hepatocite, enterocite, sau polarizata, cum este in cazul celulelor acinare pancreatice, unde RER este localizat in jumatatea bazala a celulelor, polul apical al acestora fiind ocupat de vacuolele de secreție.
|
Politica de confidentialitate |
| Copyright ©
2025 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |