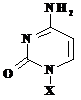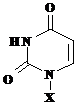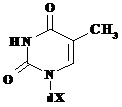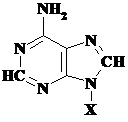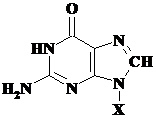Nucleotide si acizi nucleici
Nucleotidele si acizii nucleici pot fi definiti drept biomolecule care contin ca si componente esentiale in structura lor baze azotate heterociclice. Aceste biomolecule indeplinesc numeroase roluri in organismele vii, participand ca intermediari esentiali in aproape toate etapele metabolismului. Acizii nucleici sunt valuta in tranzactiile metabolice, dar si principale verigi chimice ale raspunsului celulelor la hormoni sau alti stimuli extracelulari.
Nucleotide sunt componente esentiale ale acidului deoxiribonucleic (ADN, materialul genetic al celulei) si acidului ribonucleic (ARN, cu functia de reproduce si translata informatia proteinei), substante responsabile de inmagazinarea si stocarea informatiei genetice. (Schema 5.1.) Structura fiecarei proteine este o structura programata de secventa de nucleotide dintr-un anume acid nucleic al celulei.
Schema 5.1.
5.1. Structura nucleotidelor
Toate nucleotidele au trei componente caracteristice: o baza azotata, o pentoza si o grupare fosfat. (Schema 5.2.) Lipsa partii fosfat determina structuri denumite nucleozide. Bazele azotate care intra in compozitia acizilor nucleici sunt fie de tip pirimidinic fie de tip purinic. (Schema 5.2.)
Zaharurile prezente in acizii nucleici sunt pentoze, de care se leaga resturile de baze azotate. In timp ce in ARN restul zaharic este o molecula de riboza, in ADN molecula este de tip deoxiribozic (2΄-deoxi-riboza, notarea cu "aprostof" se face pentru a deosebi notarea din ciclurile bazelor azotate cu cele din ciclurile zaharice.
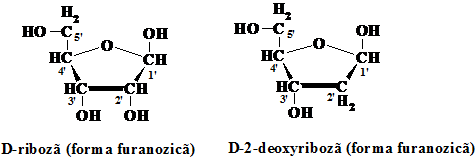
In celule se gasesc cinci baze azotate, doua cu structura purinica, adenina si guanina, si trei cu structura pirimidinica, timina (T), citozina (C) si uracilul (U). Citozina si timina se gasesc in ADN in timp ce in ARN, pe langa cantitati mici de alte baze modificate, se gaseste citozina si uracilul. (Schema 5.3.)
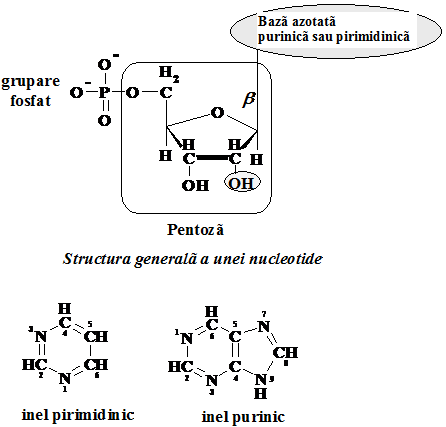
Schema 5.2. Structura nucleotidelor
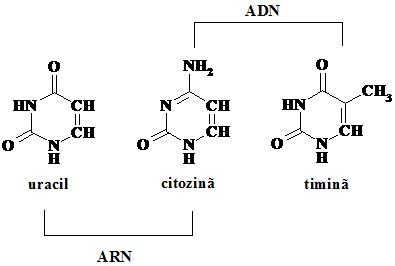
Schema 5.3.
Purinele des intalnite sunt guanina(G) si adenina (A), fiind utilizate atat de AND cat si de ARN
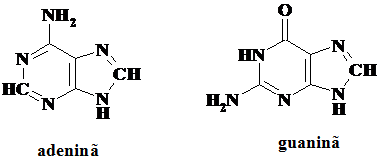
5.1.1. Proprietati ale bazelor azotate care influenteaza structura spatiala a acizilor nucleici
Purinele si pirimidinele libere au un caracter slab bazic fapt care a si condus la definirea acestora drept baze azotate. Purinele si pirimidinele din constitutia ADN sau ARN sunt molecule cu conjugare extinsa pe intreaga molecula, fapt care are consecinte deosebite asupra structurii si repartitiei electronilor si absorbtiei luminii de acizii nucleici. Aceasta conjugare puternica ofera un caracter partial de dubla legatura. Ca rezultat sistemele care au la baza inel pirimidinic sunt planare, iar inelul purinic este aproape planar. In timp ce purinele si pirimidinele libere sunt insolubile la pH fiziologic, guanina, adenina, timina, uracilul si cizozina sunt solubile datorita grupelor polare care le au.
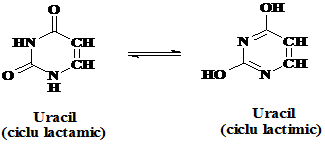
Purina si pirimidina libere poate adopta doua sau mai multe forme tautomerice (izomerice) in functie de pH. Aceasta proprietate este cunoscuta sub denumirea de tautomerie ceto-enolica. Ceto-tautomerul are un inel de tip lactam in timp ce enol-tautomerul are la baza un inel lactim. La pH fiziologic predomina forma lactam.
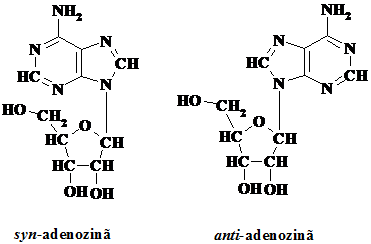
Bazele purinice si pirimidinice sunt legate in celule de un zahar, iar astfel legate sunt definite drept nucleoside. Acestea sunt legate fie la D-riboza fie la 2΄-deoxi-D-riboza prin legaturi b-N-glicozidice stabilite la azotul N9 al purinei si N1 al pirimidinei. Bazele astfel legate pot adopta doua pozitii conformationale fata de legatura N-glicozidica: syn si anti, conformatia anti fiind cea predominanta in natura.
In celule, prezenta nucleozidele este sub forma fosforilata. Aceasta forma fosforilata se defineste drept nucleotida. Cea mai obisnuita pozitie de fosforilare este gruparea hidroxil atasat de carbonul 5΄ al ribozei. Nucleotidele pot exista sub forma mono, di si trifosforilata. Forma monofosforilata este abreviata, in cazul adenozinei, AMP, iar formele di si trifosforilate ca ADP si ATP. Utilizarea acestor abrevieri presupune ca nucleotida sa fie in forma 5΄ fosforilata. Nucleotidele care se regasesc in ADN sunt diferentiate de cele din ARN prin faptul ca riboza se afla in forma ei 2΄-deoxi si pentru diferentiere forma abreviata se scrie cu un "d" in fata. Deci forma monofosforilata a adenozinei care este in ADN (deoxiadenozin-5΄-monofosfat) este presurtat scrisa ca dAMP.
Uridina nu se gaseste aproape niciodata in ADN iar timina este aproape exclusiv gasita in ADN. Exista mai putine baze azotate in ADN si ARN. Multe nucleotide modificate sunt intalnite in afara ADN sau ARN, dar indeplinesc functii biologice importante.
Toate acestea sunt prezentate in Tabelul 5.1., in care sunt reprezentate atat bazele azotate cat si nucleozidele si nucleotidele corespunzatoare.
Tabelul 5.1.
|
Formula bazei azotate |
Baza(X=H) |
Nucleozida |
Nucleotide |
|
|
Citozina, C |
Citidina, A |
Citidin monofosfat |
|
|
Uracil, U |
Uridina, U |
Uridin monofosfat |
|
|
Timina, T |
Timidina, T |
Timidin monofosfat |
|
|
Adenina, A |
Adenozina, A |
Adenozin monofosfat AMP |
|
|
Guanine, G |
Guanozina, A |
Guanozin monofosfat GMP |
5.2. Derivati reprezentativi ai nucleotidelor
5.2.1. Derivati adenozinici
Cel mai raspandit derivat adenozinic se gaseste sub forma unei structuri ciclice
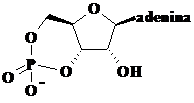 , 3΄-5΄-adenozin monofosfat ciclic (cAMP). Acest compus este un mesager secund
deosebit de important implicat in transmiterea semnalelor de la suprafata
celulelor la proteinele interne.
, 3΄-5΄-adenozin monofosfat ciclic (cAMP). Acest compus este un mesager secund
deosebit de important implicat in transmiterea semnalelor de la suprafata
celulelor la proteinele interne.
S-Adenozilmetionina este o forma activata a metioninei care serveste drept donor de grupare metilica dar si ca sursa de propilamina in sinteza poliaminelor.
5.2.2. Derivati guanozinei
La fel ca si in cazul adenozinei, un compus ciclic al guanozinei, 3΄,5΄-guanozin monofosfat ciclic (3΄,5΄-cGMP) este gasit in celule, avand rol antagonic cAMP

Cel mai important semnal asociat cGMP este cel corespunzator fotoreceptiei. Totusi, atunci cand are loc activarea rodopsinei sau a altor opsine prin absorptia unui foton se activeaza transducina care la randul ei activeaza fosfodiesteraza specifica cGMP care hidrolizeaza cGMP la GMP. Aceasta determina scaderea concentratiei efective a cGMP legat de poarta canalelor ionice, ducand la inchiderea acestora si concomitenta hiperpolarizare a moleculei.
cAMP si cGMP sunt regulatori importanti ai metabolismului celular si sunt intalniti in aproape toate celulele.
5.2.3. Derivati difosforilati si trifosforilati ai nucleotidelor
Nucleotidele monofosforilate pot lega la gruparea fosforica alte grupari de acelasi tip, rezultand compusi difosforilati respectiv trifosforilati, care prin prezenta gruparilor de tip fosfor anhidridic devin bogate energetic.
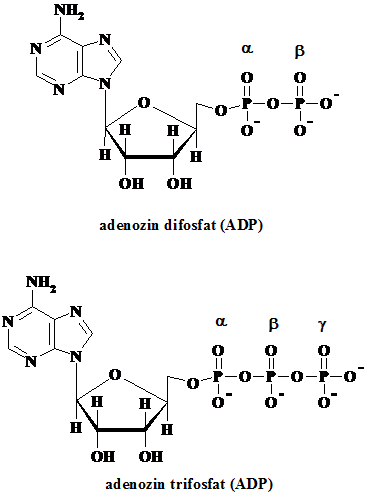
Daca se noteaza generic baza azotata prin N, atunci structurile difosfat pot fi reprezentate prin NDP, respectiv cele trifosfat prin NTP, iar deoxinucleozidele corespunzatoare se vor nota cu dNDP si dNTP. Notarea atomilor de fosfor se face cu litere grecesti incepand cu atomul de fosfor legat prin punte de oxigen de zahar. Aceste structuri di- si trifosforilate se gasesc ca molecule libere in celule.
Dintre caracteristicile lor evidentiem faptul ca la deprotonare totala pot elibera trei ioni de hidrogen (H+), in cazul NDP si patru protoni in cazul NTP, fiind astfel acizi poliprotici. Pe de alta parte fiecare tip in parte are rolul sau unic in celula: ATP ca banca energetica a celulei; GTP reprezinta principala sursa energetica pentru sinteza proteinelor; CTP este metabolit esential in sinteza fosfolipidelor; UTP folosit la activarea intermediarilor in biosinteza carbohidratilor. De remarcat ca in toate cazurile, baza azotata este cea care contine informatia ce directioneaza spre functia specifica, deoarece ea este cea care difera intre aceste biomolecule.
5.3. Oligonucleotide
Oligonucleoditele reprezinta baza structurala a acizilor ARN si ADN, reprezentand polimeri ai acizilor nucleici. Sunt sintetizati cu ajutorul enzimelor polimeraze si diferitelor NTP sau dNTP. Caracteristic acestora este legarea diesterica, printr-o grupa fosforica, la pozitiile 3' si 5' ale zaharurilor acizilor nucleici. Deoarece pozitia initiala a gruparii monofosfat este 5', iar legarea se face la un carbon 3' al celuilalt acid nucleic, noua legatura formata mai este descrisa drept 3' 5' fosfodiesterica, indicandu-se astfel directia cresterii macromoleculei.
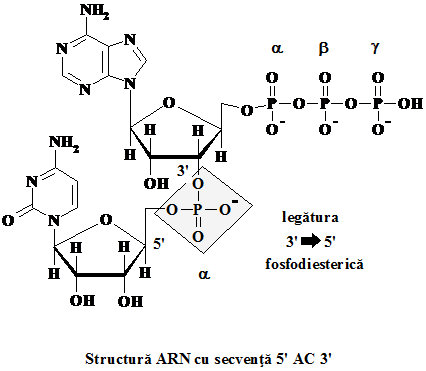
In consecinta, intr-o polinucleotida capatul 5' reprezinta "startul" respectiv capatul 3' este "sfarsitul". De subliniat ca de obicei capatul 5' este monofosfat, iar 3' un hidroxil, fara insa ca aceste sa fie absolut configurativ obligatoriu.
5.4. Structura acizilor dezoxiribonucleici
Acizii dezoxiriboinucleici sunt materialul genetic al cromozomilor, fiind localizati mai ales in nucleul celulelor. Structura lor este una polimerica, caracteristica secventiala fiecarei specii, alcatuita din mii sau chiar milioane de monucluceotide legate prin intermediul legaturii 3' 5' fosfodiesterica prezentata anterior.
Structura stabilita de James Watson si Francis Crick, prin spectroscopie de raze X a cristalelor de ADN, sugereaza un model tip "helix" (elice). Modelul prezinta doua fasii antipararele impletite una in jurul celeilalte, fiind stabilizate prin legaturi de hidrogen. Modelul mai arata ca bazele azotate se gasesc in interiorul elicei, aproape perpendiculare pe axul elicii.
Pe de alta parte, determinarile experimentale au stabilit ca in oricare molecula de ADN concentratia adeninei (A) este egala cu cea a timinei (T) iar a citidinei (C) egala cu cea a guanidinei (G). Acestea au sugerat, asa cum a fost de fapt si stabilit, formarea de perechi doar intre acesti acizi nucleici.

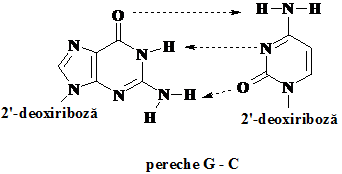
Deoarece perechea G -C contine trei legaturi de hidrogen comparative cu cele doua ale perechii A -T, aceasta este o pereche mai stabila.
Orientarea antiparalela consta in faptul ca in elice una din macromolecule este orientata directia 5', iar cealalta in 5' 3'. Gruparile fosfat sunt situate in afara elicei iar perechile bazele azotate in interiorul acestora.
![]()
Diametrul elicei este de 20 Å, cu o perioada de identitate de 34 Å. Cele doua policatene sunt rasucite orientativ spre dreapta, o spira avand cate 10 nucleotide.
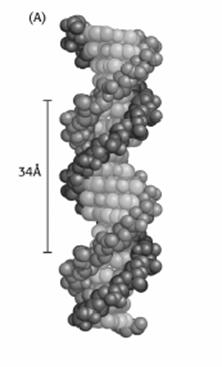
Figura 5.1. Elice de ADN forma A
Elicea ADN poate lua diferite forme, in functie de continutul secventional, conditiile ionice ale separarii cristaline. Forma B este predominanta in cazul unei inalte hidratari si a unei concentratii ionice mici. Forma Z reprezinta o modificare in sensul rotirii spre stanga a policatenelor elicei. Parametri caracteristici celor trei forme ale elicei ADN sunt prezentati in tabelul 5.2.
Tabelul 5.2. Parameterii eliciilor de ADN
|
Parametru |
Forma A |
Forma B |
Forma Z |
|
Directia de rotatie a elicei |
dreapta |
dreapta |
stanga |
|
Perioada de identitate |
12 perechi baze |
||
|
Orientarea legaturii N-glicozidice |
Anti |
Anti |
Anti pentru Piridine, Syn pentru Purine |
5.5. Acizii ribonucleici
Acizii ribonucleici sunt prezenti in aproape toate partile unei celule. Se deosebesc trei tipuri mari de ARN.
ARN Ribozomial (r ARN) apare in ribozomii citoplasmatici, fiind o molecula inalt polimerizata cu masa moleculara de 5x105 - 1x106.
ARN Mesager (m ARN), cu masa moleculara cuprinsa intre 25x103 si 3x105 si sintetizat in nucleu in procesul de transcriptie, este purtatorul informatiei, constituind practic o matrita in citoplasma.
ARN de Transfer (t ARN) este constituit din policatene mai mici avand rol de transfer in procesele metabolice.
|
Politica de confidentialitate |
| Copyright ©
2025 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |