
STRUCTURA PRIMARA A ACIZILOR NUCLEICI
Asemenea poliglucidelor si proteinelor, acesti polimeri se caracterizeaza printr-o structura covalenta primara si prin aranjarea spatiala a acesteia intr-o ierarhie pe care o vom discuta in capitolele urmatoare.
Structura primara a acizilor nucleici (AN) se refera la secventa inlantuirii monomerilor nucleozid 5'-fosfat (NMP), legaturile dintre acestia si marimea polimerilor obtinuti.
Caracteristicile de compozitie a structurii primare
Nucleotidele
Cele doua tipuri de acizi nucleici se deosebesc prin doua caracteristici calitative fundamentale (Tabelul 2.1
1) prezenta unei pentoze diferite (riboza sau deoxiriboza); tipul de glucid da numele celor doua tipuri de acizi nucleici deoarece nu exista structuri naturale mixte ADN/ARN in afara de hibrizii care apar tranzitoriu in timpul replicarii si transcriptiei;
2) diferente privind cele patru baze care intra in constitutia ADN si ARN, una dintre pirimidine fiind diferita. O distinctie mai nuantata este data de bazele si nucleotidele minore discutate in capitolul anterior.
Tabel 2. Constituentii ARN si ADN
|
Acidul nucleic |
Pentoza |
NMP majore |
NMP minore |
|
|
Pyr |
Pur | |||
|
ARN |
D-riboza |
U C |
A G |
foarte diverse ARNt |
|
ADN |
2'-deoxi-D-riboza |
dT dC |
dA dG |
derivati metilati ai A si C |
Cu toate ca aceste diferente par minime ele au consecinte spectaculoase si sunt suficiente pentru a imparti acizii nucleici in doua categorii majore de molecule destul de asemanatoare pentru a se recunoaste dar care au functii diferite. Astfel, prezenta hidroxilului in pozitia 2' a ribozei determina:
a) imposibilitatea catenelor de ARN de a forma structuri dublu catenare pe distante mari;
b) o reactivitate particulara a ARN;
c) centre suplimentare de interactiune prin legaturi de hidrogen.
O alta diferenta de compozitie consta in faptul ca ADN se caracterizeaza prin egalitati si raporturi purine/pirimidine specifice pe care nu le regasim in ARN.
Marimea polimerilor nucleici
In privinta lungimii polimerilor nucleici gama acoperita este foarte mare. De fapt, trebuie sa subliniem ca este luat in discutie grupul care cuprinde macromoleculele naturale cele mai mari. Marimea acizilor nucleici se exprima prin trei caracteristici (tipuri de unitati): i) lungime; ii) masa moleculara in Daltoni (Da); iii) numar de nucleotide sau de baze ale acestora pentru monocatene (b) si in perechi de baze (pb) daca molecula este constituita din doua catene asociate. Multiplul cel mai folosit este cel de kilobaze sau kilo-perechi de baze ( 1000 b sau 1000 pb).
Numarul nucleotidelor din structura ARN variaza de la mai multe zeci de baze la mai multe mii de baze.
ARN ribozomal (ARNr) de la 100 la 5 000 b
ARN de transfer (ARNt) 75 - 90 b
ARN mesager (ARNm) numarul de baze este in functie de gena copiata
Marimea ADN variaza in functie de regn si specie. Aceasta acopera toata gama de la 5 000 la mai mult de un milion de perechi de baze. Masa moleculara medie a unei perechi de baze este de 660 Da, iar distanta intre platourile a doua baze este de 0, 34 nm in conformitate cu caracteristicile geometrice ale dublei catene de ADN. Aceste caracteristici dau o echivalenta foarte utila in practica: 2x106 Da se pot aproxima cu
000 pb si cu mm
In tabelul 2.2. sunt prezentate tipuri de ADN de provenienta diferita. Marimea acestora este comparata cu colagenul care este o molecula proteica gigant.
Metode de determinare a marimii acizilor nucleici
 Metodele
de determinare a dimensiunilor acestor macromolecule se bazeaza pe
diferite caracteristici. Marimea poate fi determinata prin mai multe
metode:
Metodele
de determinare a dimensiunilor acestor macromolecule se bazeaza pe
diferite caracteristici. Marimea poate fi determinata prin mai multe
metode:
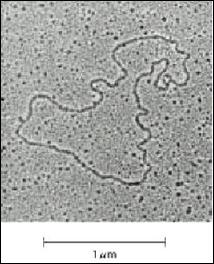
Microscopie electronica: datorita dimensiunilor lor acizii nucleici pot vizualizati prin aceasta tehnica. Metoda se potriveste foarte bine moleculelor de ADN de marime medie (cateva mii la 200 000 pb) (Figura 2.1
Figura 2. Imagine electrono-microscopica a unei molecule de ADN dublu catenara circular inchisa (plasmida)
Electroforeza in gel: este o tehnica de separare a moleculelor in functie de masa lor moleculara. Conditiile de densitate de sarcina asemanatoare sunt realizate in mod natural, de catre scheletul oza-fosfat. De asemenea, dependenta logaritmica, distanta de migrare in functie de marime, este bine respectata. Pentru separarea moleculelor de acizi nucleici de diferite dimensiuni sunt folosite geluri de agaroza cu porozitati variate (Figura 2.2). Pentru facilitarea estimarilor cantitative exista colectii de polimeri "marcheri de masa moleculara" utilizati pentru etalonarea separarilor. De altfel, aceasta metoda de separare este omniprezenta in manipularile de biologie moleculara. Electroforeza se poate realiza si in geluri de poliacrilamida pentru situatiile in care este necesara separarea moleculelor polinucleotidice mici sau oligonucleotidelor. Prin electroforeza in gel de poliacrilamida este posibila identificarea unor fragmente care difera numai printr-un singur nucleotid. Din acest motiv tehnica este foarte utila in secventializarea acizilor nucleici si in determinarea regiunilor de polimorfism normal sau patologic determinat de existenta unor mutatii.
Tabel 2. Marimea unor molecule de ADN din diferite surse, comparata cu cea a unei proteine fibroase, colagenul, si celulele procariote si eucariote.
|
ADN/ particule/ organisme |
n de baze (kb) |
Masa (MDa) |
Lungime (nm) |
f (nm) |
numar de cromozomi |
|
Virusuri ADN SV40 fag FX174 fag l |
1,7x10-3 (1,7mm) 1,8x10-3 (1,8 mm) 17x10-3 (17 mm) | ||||
|
Procariote E. coli | |||||
|
Eucariote Drosofila Om |
2x4 2x23 |
||||
|
Colagen |
3x1000 aminoacizi |
0,3x10-3 (0,3 mm) | |||
|
Celule Bacteria E. coli Mamifere |
2x10-3 (0,5x2 mm) aprox 10x10-3 (10 mm) | ||||
SV40: virus simian; marimea exacta a ADN este de 5243 pb. Bacteriofagii folositi ca exemplu au drept gazda Escherichia coli. ADN al fagului FX174 este monocatenar, dar in celulele infectate se imperecheaza sub forma de dubla catena. In tabel sunt date dimensiunile celei de a doua forme. La drosofila este data marimea celui mai mare ADN, este vorba de cromozomul gigant. La om este data marimea ADN/cromozom.
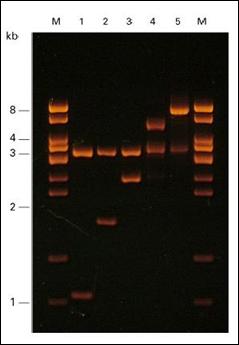 Figura 2.2 Imagine de
separare electroforetica a ADN in gel de agaroza: M, marcheri de
masa moleculara; 1, 2, 3, 4 si 5, ADN plasmidial scindat cu
enzime de restrictie
Figura 2.2 Imagine de
separare electroforetica a ADN in gel de agaroza: M, marcheri de
masa moleculara; 1, 2, 3, 4 si 5, ADN plasmidial scindat cu
enzime de restrictie
Moleculele ADN de marime mare sunt analizate si prin
a) metoda clasica de ultracentrifugare cu toate ca fortele de frecare existente intre aceste molecule limiteaza campul aplicatiilor (molecule de pana la 1,5 x 106 pb); Prin aceasta tehnica pot fi separate, in gradient de densitate de clorura de cesiu, urmatoarele: i) fragmente de ADN care au un continut diferit in guanina si citozina (Figura 2.3a); ii) ADN genomic bacterian; iii) ADN plasmidial; iv) ARN total, etc. Diferitele tipuri de ARN ribozomal de la eucariote si procariote poate fi separat foarte simplu prin ultracentrifugare in gradient de zaharoza (Figura 2.3b
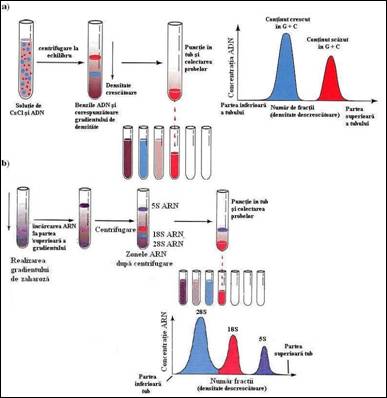
Figura 2. Separarea acizilor nucleici prin ultracentrifugare: a) Separare ADN prin ultracentrifugare in gradient de CsCl; b) separare ARN prin ultracentrifugare in gradient de zaharoza
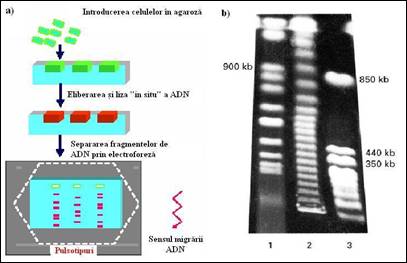
Figura 2. Electroforeza in camp pulsatoriu. a) principiu de separare; b) electroforegrama obtinuta in urma separarii unor fragmente mari de ADN. Vizualizarea a fost realizata in urma colorarii cu bromura de etidium.
b) proprietatile hidrodinamice ale solutiilor de ADN: i) birefringenta de curgere, ii) vascozitate, etc.
c) electroforeza in camp pulsatoriu (sau alternant). Prin aceasta tehnica care a fost imaginata la inceputul anilor 1980, doua campuri electrice se aplica alternativ pe doua directii diferite. Principul de separare se bazeaza pe viteza diferita de reorientare a moleculelor in momentul alternarii celor doua campuri electrice. Ele impun moleculelor de ADN etape de reorientare diferite inainte de migrarea lor efectiva. Aceste etape de reorientare sunt dependente de marimea moleculelor. Separarea este eficace si utilizabila pentru molecule de ADN mai mari de 10 000 kb (Figura 2.4
Polimerizarea nucleotidica
Acizii nucleici sunt, asemeni proteinelor, polimeri neramificati care pot fi liniari sau circulari. Demonstrata de Levene in 1925 si confirmata dupa 1950 de catre Todd, inlantuirea covalenta a nucleotidelor este de tip ester. Depolimerizarea se poate realiza prin hidroliza de diferite tipuri (acida, bazica, enzimatica) si este destul de folosita in tehnicile de biologie moleculara.
Legatura fosfodiester
Indiferent de tipul de acid nucleic (ribonucleic sau deoxiribonucleic), reactia de asamblare polinucleotidica este aceeasi: hidroxilul din pozitia 3' al unui nucleozid monofosfat este esterificat de catre 5'-fosfatul din structura nucleozid trifosfatului care participa la formarea legaturii (Figura 2.5). Faptul ca acizii nucleici sunt polimeri fosfodiesterici ai NMP are urmatoarele consecinte directe:
a) asemeni polipeptidelor, catena este orientata (vectorizata). Sensul conventional 5'-3' a fost adoptat pentru lectura si scrierea (de la stanga la dreapta) polinucleotidelor ca si pentru reprezentarile prescurtate si simbolice (Figura 2.6
b) bazele fixate regulat in functie de o secventa specifica speciei moleculare, sunt purtate de un schelet covalent pentoza-fosfat. Cu toate ca imaginea este asemanatoare cu cea a scheletului polipeptidic purtator al catenelor laterale ale aminoacizilor, exista o serie de diferente fata de acesta: i) scheletul acizilor nucleici este un polianion cu o sarcina pe unitate care are o densitate totala de sarcina negativa foarte mare; ii) varietatea bazelor este redusa. Analizam alternanta a patru baze fata de alternanta a 20 de aminoacizi. Consecintele acestor diferente sunt foarte importante pentru sistemele de informatie genetica. Intelegem astfel, de ce pentru codificarea aminoacizilor de catre secventele nucleotidice nu este nevoie numai de o simpla echivalenta baza/aminoacid. Este necesar un limbaj ternar, format din ansambluri de trei baze adiacente a caror asociere determina existenta a 64 de combinatii posibile, care stau la baza codului genetic.
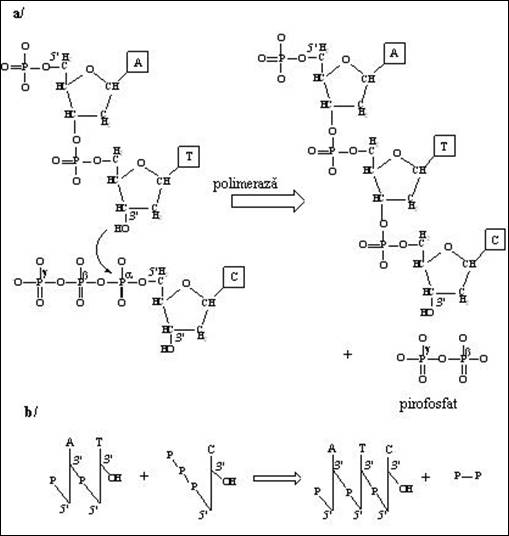
Figura 2. 5. Sinteza unei monocatene de ADN: a) formarea legaturii fosfodiester intre un dinucleotid si deoxiadenozin citidin trifosfat; b) reprezentarea schematica a reactiei de sinteza cu marcarea orientarii 5'-3' a catenei in formare
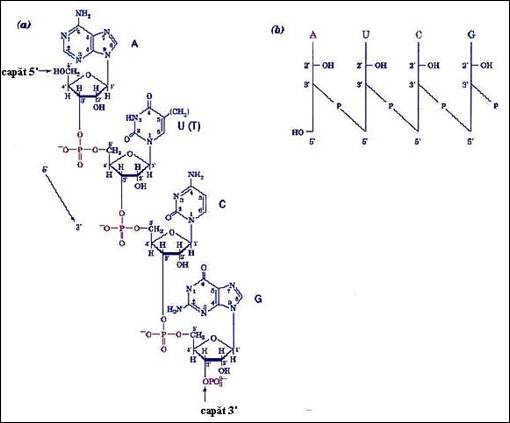
Figura 2.6. Structura unei molecule ARN: a) evidentierea inlantuirii nucleotidelor si vectorizarea catenei; b) reprezentarile prescurtata si simbolica.
Hidroliza acizilor nucleici
Degradarea unui polinucleotid poate sa se refere la: i) legatura fosfodiester atunci cand se produce o depolimerizare; ii) unitatile nucleotidice daca se rupe legatura glicozidica si se degradeaza componentele. In ambele cazuri hidroliza poate fi de natura chimica sau enzimatica.
Hidroliza chimica a acizilor nucleici
Desi sunt printre cele mai stabile molecule, in conditii fiziologice celulare, acizii nucleici prezinta totusi, rezistente diferite la conditiile de pH si de temperatura la care se realizeaza diferitele tipuri de hidroliza, in vitro (Tabel 2.3
Tratamentul acid afecteaza la fel ARN si ADN
i) sunt necesare conditii drastice pentru a degrada scheletul pentozo-fosfat (incalzire, acizi concentrati). In aceste conditii celelalte legaturi din structura nu rezista si obtinem un amestec de fosfati, oze si baze. Acest protocol, urmat de o fractionare cromatografica, a fost una dintre tehnicile care au permis analiza compozitiei acizilor nucleici;
i) in conditii relativ blande (pH 4,0), se vor rupe numai legaturile N-glicozidice cu purinele care sunt cele mai fragile. In acest caz, se obtine un derivat de acid nucleic apurinic care prezinta interes pentru anumite analize.
Comportamentul ARN si ADN la hidroliza bazica este foarte diferit:
i) ADN rezista la valori extreme de pH bazic. La pH 13,0 si 370C se inregistreaza aproximativ zece scindari pe ora si pentru milion de punti fosfo-diester. Cu toate acestea, puntile fosfodiester din ADN devin sensibile la hidroliza alcalina daca bazele sunt inlaturate sau degradate. Este cazul ADN apurinic. Aceasta proprietate este utilizata in tehnica de secventializare chimica.
ii) ARN este total hidrolizat in ribonucleotidele componente in cateva minute, la 370C si pH 11,0. Inca de la pH 8,0, se constata inceperea unei degradari semnificative. Comportamentul ARN la hidroliza alcalina explica obtinerea unui amestec de nucleozide 2' si 3' fosfat in loc de 5' fosfat, asa cum era de asteptat.
Tabel 2. Produsii de degradare chimica a acizilor nucleici
|
Conditii |
Produsi de hidroliza |
|
|
ARN |
ADN |
|
|
Acide pH 1,0 incalzire pH 4,0 |
pentoze + fosfati + baze baze purinice + AN apurinici |
|
|
Alcaline |
NMP (2' sau 3') | |
Diferenta dintre reactivitatea ARN si inertia ADN este data de existenta gruparii hidroxil din pozitia 2'. De fapt, aceasta grupare autorizeaza o cataliza bazica hidrolizei puntilor fosfodiester schematizata in figura 2.7. Deprotonarea sa in conditii alcaline produce un anion alcoxid, puternic nucleofil care ataca gruparea fosfo-diester adiacenta. Absenta hidroxilului in 2' din scheletul ADN este un factor determinant al stabilitatii sale chimice in conditii celulare, normale sau agresive. Astfel intelegem esenta selectionarii evolutive a ADN, ca suport al informatiei genetice, datorita rezistentei sale. Aceasta molecula trebuie sa suporte operatii de duplicare si de copiere care se desfasoara pe toata durata de viata a organismului si sa asigure transmiterea informatiilor la descendenti.
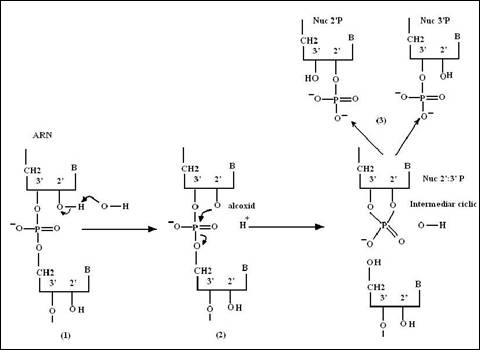
Figura 2. Schema mecanismului de hidroliza ARN prin cataliza bazica: 1. Hidroxilul din 2' este deprotonat de catre ionul hidroxid; 2. Alcoxidul, nucleofil, ataca gruparea fosfodiester cu formarea unui diester ciclic 2': 3'; 3. Un al doilea hidroxid provoaca hidroliza uneia sau alteia dintre legaturile ester: se obtin izomeri 2' si 3' fosfat.
Scindarea enzimatica
Enzimele care catalizeaza hidroliza legaturii fosfodiester din structura acizilor nucleici, fosfodiesterazele, sunt numite si nucleaze. Acestea sunt prezente in toate celulele si sunt foarte utile in metabolismul acizilor nucleici si proteinelor. O parte dintre a ceste nucleaze sunt secretate de catre pancreasul exocrin, in intestin, si participa direct la digestie hranei. Aceste enzime prezinta niveluri de specificitate comparabile cu cele ale peptidazelor si proteazelor. Ele se pot clasifica in functie de:
a) modul lor de atac al catenei: i) exonucleaze, enzimele care actioneaza la nivelul extremitatii catenelor; ii) endonucleaze, enzimele care ataca in interiorul catenelor;
b) specificitatea lor fata de substrat: i) pentru tipul de acid nucleic: ribonucleaze pentru ARN, deoxiribonucleaze pentru ADN si nucleaze pentru ambele; ii) pentru tipul de structura: mono- sau dublu catenara;
c) specificitatea lor de recunoastere a situsurilor: i) specificitate pentru baze; ii) specificitate pentru secvente;
d) tipul de rupere a legaturii fosfodiester: i) exonucleazele si endonucleazele de tip d scindeaza extremitatea 5' si duc la formarea de 3'NMP; ii) exonucleazele si endonucleazele de tip p scindeaza extremitatea 3' si determina formarea 5'NMP (Figura 2.8
In tabelul 2.4 sunt prezentate principale nucleaze si caracteristicile acestora. O serie de endonucleaze cu specificitate mare nu sunt trecute in acest tabel. Acestea fac parte din categoria enzimelor de restrictie, endonucleaze capabile sa scindeze intr-un mod, bine definit si reproductibil, moleculele de ADN dublu catenar indiferent de originea lor.
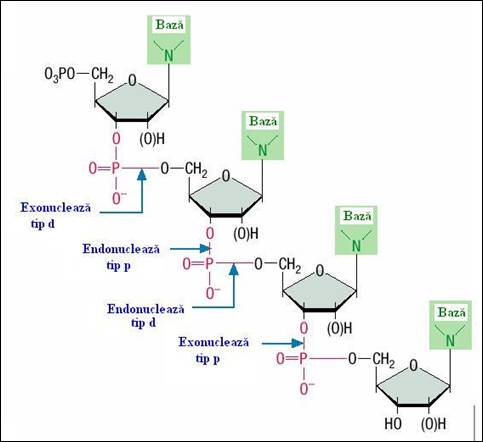
Figura 2.8. Tipuri de taieturi la nivelul legaturilor: exonuclea-zele si endonucleazele de tip d scindeaza extre-mitatea 5' si duc la formarea de 3'NMP; exonucleazele si endo-nucleazele de tip p scindeaza extremitatea 3' si determina formarea 5'NMP (vezi tabelul 2.4)
Tabel 2. Exemple de nucleaze reprezentative si diferitele lor caracteristici.
|
Nucleaze |
Substrate1 |
Taietura tip2; specificitate |
Produsi |
|
Exonucleaze |
|||
|
fosfodiesteraza (venin de sarpe) |
ARN, ADN s |
p extremitatea 3' |
5'-NMP |
|
fosfodiesteraza (splina) |
ARN, ADN s |
d extremitatea 5' |
3'-NMP |
|
exonucleaza I (E. coli) |
AND |
p extremitatea 3' |
5'-NMP |
|
exonucleaza III (E. coli) |
AND |
p extremitatea 3' |
5'NMP+ADNs |
|
Endonucleaze |
|||
|
endonucleaza S1 (Aspergillus oryzae) |
ARN,ADN s,d |
p aleatoare |
5'-NMP +ON in 5' P |
|
ribonucleaza T1 (Aspergillus oryzae) |
ARN s |
p G/N |
5'-NMP +ON in 3' P |
|
ribonucleaza A (pancreas) |
ARN s |
d-Pyr/N |
3'-NMP +ON in 3' P |
|
Deoxiribonucleaza II (timus) |
ADN s |
d sPyr/dPur |
ON in 3' P |
s: monocatenar (single strand); ON: oligonucleotide. tip de taietura p sau d: vezi figura 2.8
Descoperirea enzimelor de restrictie, in 1970, de catre W. Harber, H. Smith si D. Nathans, a insemnat un pas gigantic pentru evolutia tehnicilor de biologie moleculara. Acestea au permis inlaturarea principalului obstacol tehnologic privind manipularea ADN, deoarece pot reduce intr-o maniera reproductibila, un genom intreg la o serie de fragmente caracteristice pentru o specie data. Astfel, genele sau parti ale genelor devin entitati fizice izolabile. Aceste enzime sunt responsabile pentru fenomenul de restrictie de unde si numele lor. In figura 2.9 sunt prezentate cateva dintre enzimele de restrictie cele mai uzuale impreuna cu motivele recunoscute de ele. Enzimele recunosc si taie secvente de baze cu structuri caracteristice de pe catena ADN, numite palindroame, a caror lungime variaza intre 4 si 10 pb. Se remarca axa de simetrie de ordinul 2 pe care o prezinta secventele ADN pe care aceste enzime le recunosc si scindeaza. Se observa ca secventa de pe una din catene reprezinta palindromul secventei celeilalte catene. Pe cele doua catene exista acelasi mesaj care poate fi citit la fel in cele doua sensuri (de tip RADAR sau ROTOR). Deci, situsul de clivare are o pozitie simetrica in raport cu axa. In urma actiunii enzimelor de restrictie, se pot obtine capete coezive sau capete drepte pentru fragmentele eliberate, in functie de pozitia situsurilor de scindare. (Figura 2.10
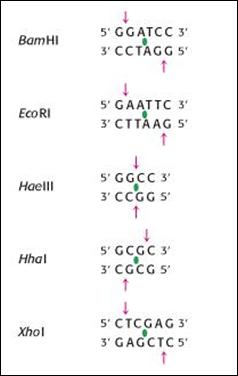
Au fost izolate si purificate mii de enzime de restrictie. Din fericire foarte curand dupa descoperirea lor s-a utilizat o nomenclatura adecvata. Prima litera este majuscula si reprezinta initiala speciei bacteriene din care a fost extrasa endonucleaza. Urmatoarele doua litere sunt minuscule si corespund genului bacteriei de unde enzima este extrasa. Aceste trei litere sunt urmate de o cifra romana care reprezinta numarul de ordine al descoperirii enzimei in acelasi tip de bacterie. Daca este necesar se mai adauga o litera majuscula reprezentand identificarea susei bacteriene.
Figura 2.9. Exemple de enzime de restrictie si situsurile recunoscute de acestea
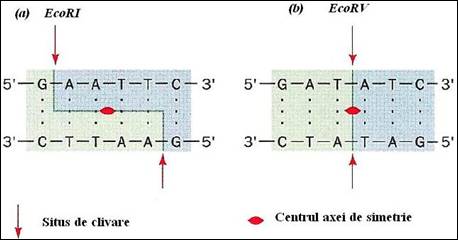
Figura 2.10. Exemple de enzime de restrictie si palindroamele recunoscute de acestea cu marcarea situsurilor de clivare si a axei de simetrie: a) obtinerea de capete coezive; b) obtinerea de capete drepte (dupa Biochemistry, Voet&Voet, 1995)
Enzimele de restrictie sunt clasificate in trei categorii, dar numai cele de tip II sunt utile in cercetarile de biologie moleculara deoarece scindeaza catenele de ADN chiar la nivelul situsurilor specifice pe care le recunosc. Doua enzime de restrictie care recunosc aceeasi secventa sunt numite izoschizomere
Numarul de situsuri recunoscute si taiate de anumite enzime este evident corelat cu structura ADN. In tabelul 2.5 este prezentat un exemplu de actiune diferentiata a enzimelor de restrictie asupra ADN viral din diferite surse. Scindarea diferentiata reprezinta o modalitate importanta de a obtine marcheri de masa moleculara utilizati in tehnicile de biologie moleculara.
Tabel 2. Numarul situsurilor de taiere prezente la nivelul ADN din trei virusuri pentru trei endonucleaze comune
|
Endonucleaza (sursa) |
Numar de situsuri de taiere |
||
|
Virus SV40 |
fag FX174 |
fag l |
|
|
EcoRI (E coli RY | |||
|
BamHI (Bacillus amyloliquefaciens H | |||
|
HaeIII (Haemophilus aegypticus) |
>50 |
||
Purificarea acizilor nucleici si evaluarea cantitativa a acizilor nucleici
Un procedeu clasic de purificare consta in tratarea solutiei apoase de acid nucleic cu solutie fenolica care denatureaza proteinele. De obicei este folosit amestecul fenol-cloroform care este destul de eficient. Dupa separarea fazelor prin centrifugare, este recuperata faza apoasa care constituie supernatantul si care contine acizii nucleici. Urmele de fenol (care ar putea sa inhibe activitatea enzimelor din etapele urmatoare) sunt eliminate printr-un nou tratament cu cloroform/alcool izoamilic. Daca acizii nucleici provin din extracte celulare, proteinele sunt in prealabil hidrolizate cu ajutorul unei enzime proteolitice numita proteinaza K.
In ultimii ani au fost puse la punct procedee de purificare pe baza de rasini. Reactivii sunt livrati sub forma de "kituri" gata de intrebuintare simplificand mult procesul de purificare si crescand adesea randamentul acestuia.
Metode spectrofotometrice
Determinarea absorbtiei in ultraviolet cu ajutorul unui spectrofotometru permite verificarea puritatii unei solutii de ADN sau ARN. In urma determinarii densitatii optice se va obtine un pic la 260 nm. Pentru estimarea puritatii este necesara calcularea raportului densitatilor optice la 260 nm/280 nm care, in mod normal, trebuie sa fie apropiat de valoarea 1,8-2,0. In cazul ADN, un raport mai ridicat indica o contaminare cu ARN. Daca exista o contaminare cu proteine (280 nm) sau fenol (270 nm), raportul va fi net inferior valorii de 1,8. Daca ADN si ARN sunt puri, cantitatea poate fi apreciata astfel:
unitatea DO (260 nm) = 50 ng/ml pentru ADN dublu-catenar
unitatea DO (260 nm) = 33 ng/ml pentru ADN monocatenar
unitatea DO (260 nm) = 40 ng/ml pentru ARN
Utilizarea de mini-gelulri
Daca nu dispunem de ADN in cantitate suficienta sau de sisteme miniaturizate care sa permita masurarea densitatii optice la 260 nm, se poate realiza o estimare semi-cantitativa utilizand mini-geluri de agaroza. Dupa realizarea separarii electroforetice, se realizeaza colorarea cu bromura de etidium urmata de evaluarea intensitatii benzilor obtinute dupa expunerea la UV. Estimarea semi-cantitativa se realizeaza prin compararea rezultatelor obtinute cu migrarea unor cantitati cunoscute de ADN standard folosite pentru etalonare.
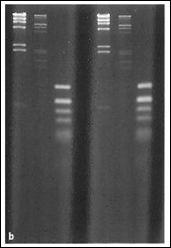
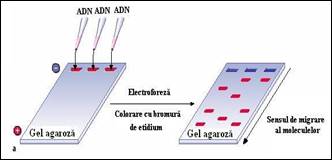
Figura 2. Separare electroforetica pe mini-geluri utilizata pentru estimarea semi-cantitativa a fragmentelor de acizi nucleici separate: a) principiu de separare; b) vizualizarea mini-gelului dupa tratament cu bromura de etidium
|
Politica de confidentialitate |
| Copyright ©
2025 - Toate drepturile rezervate. Toate documentele au caracter informativ cu scop educational. |
Personaje din literatura |
| Baltagul caracterizarea personajelor |
| Caracterizare Alexandru Lapusneanul |
| Caracterizarea lui Gavilescu |
| Caracterizarea personajelor negative din basmul |
Tehnica si mecanica |
| Cuplaje - definitii. notatii. exemple. repere istorice. |
| Actionare macara |
| Reprezentarea si cotarea filetelor |
Geografie |
| Turismul pe terra |
| Vulcanii Și mediul |
| Padurile pe terra si industrializarea lemnului |
| Termeni si conditii |
| Contact |
| Creeaza si tu |